

An essential role of Glu-243 and His-239 in the phosphotransfer reaction catalyzed by pyruvate dehydrogenase kinase
- PMID: 11278487
- PMCID: PMC2144735
- DOI: 10.1074/jbc.M009327200
An essential role of Glu-243 and His-239 in the phosphotransfer reaction catalyzed by pyruvate dehydrogenase kinase
Abstract
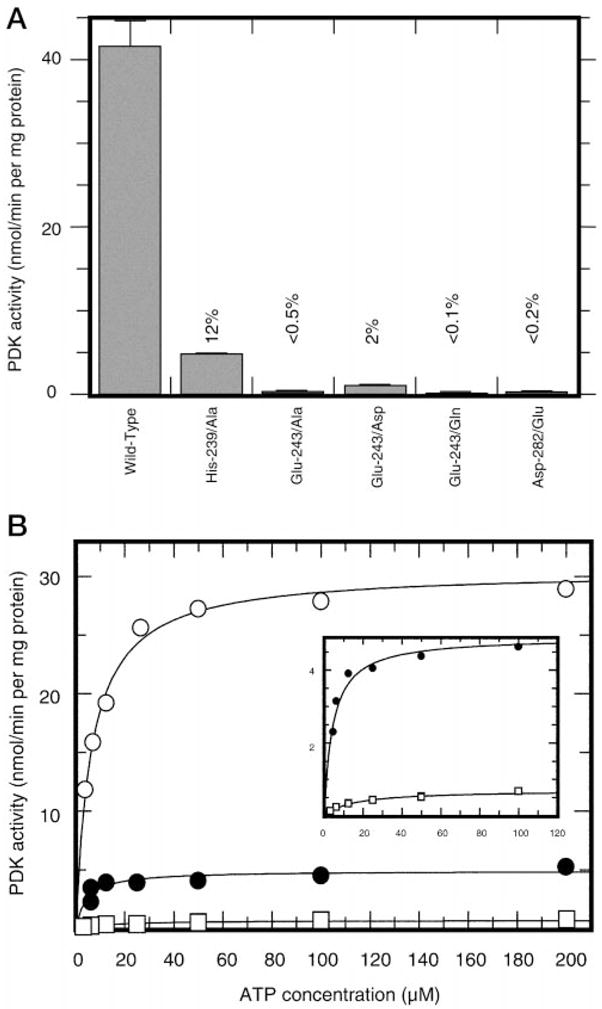
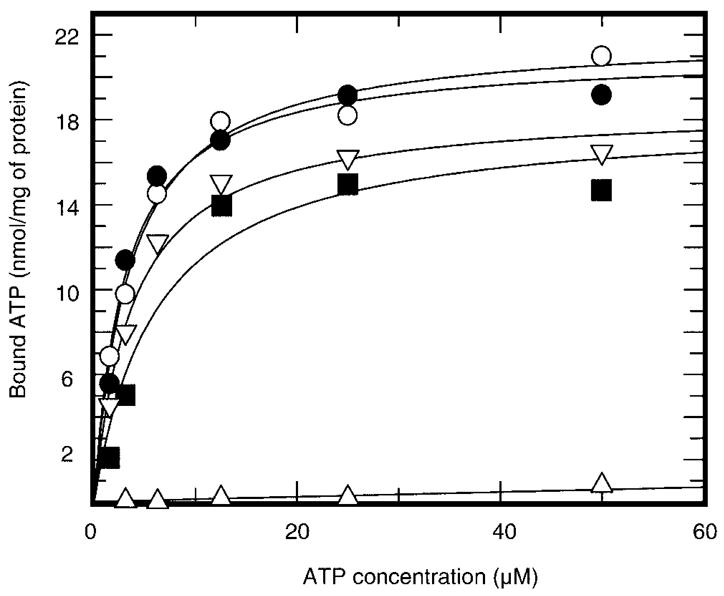
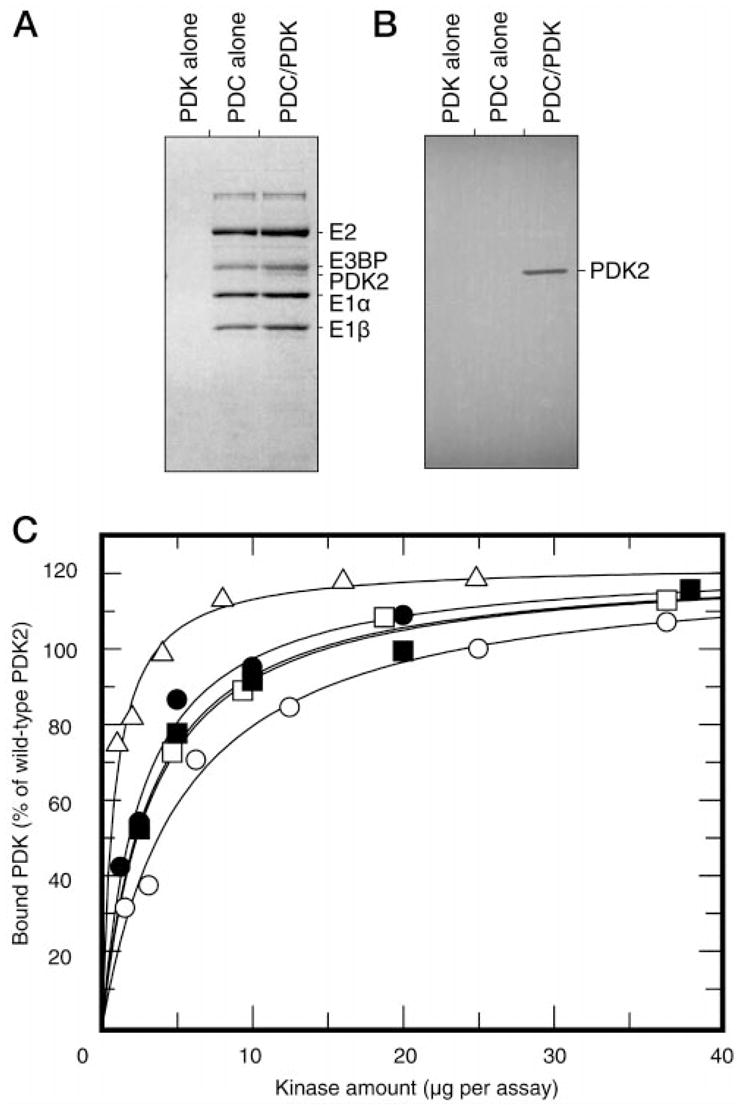
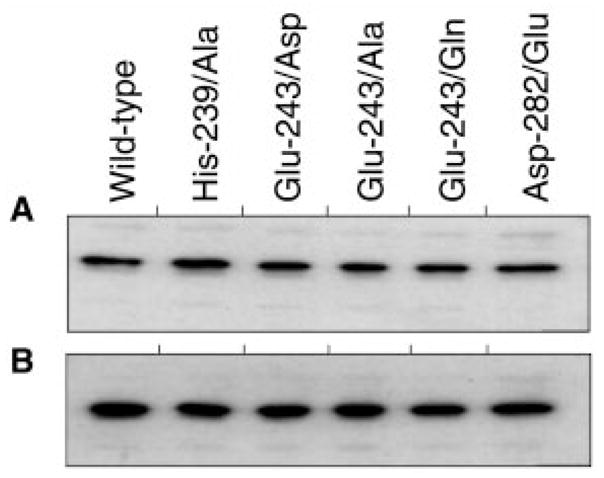
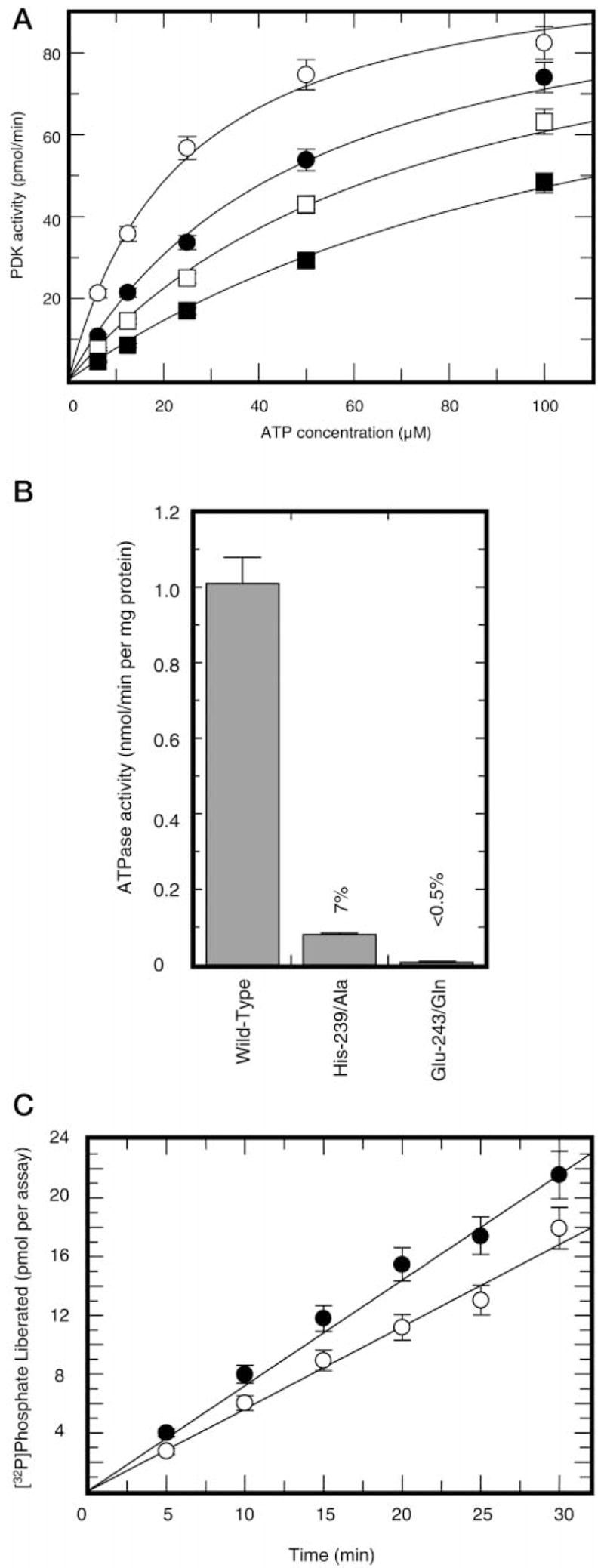
This study was undertaken to examine the mechanistic significance of two highly conserved residues positioned in the active site of pyruvate dehydrogenase kinase, Glu-243 and His-239. We used site-directed mutagenesis to convert Glu-243 to Ala, Asp, or Gln and His-239 to Ala. The resulting mutant kinases demonstrated a greatly reduced capacity for phosphorylation of pyruvate dehydrogenase. The Glu-243 to Asp mutant had approximately 2% residual activity, whereas the Glu-243 to Ala or Gln mutants exhibited less than 0.5 and 0.1% residual activity, respectively. Activity of the His-239 to Ala mutant was decreased by approximately 90%. Active-site titration with [alpha-(32)P]ATP revealed that neither Glu-243 nor His-239 mutations affected nucleotide binding. All mutant kinases showed similar or even somewhat greater affinity than the wild-type kinase toward the protein substrate, pyruvate dehydrogenase complex. Furthermore, neither of the mutations affected the inter-subunit interactions. Finally, pyruvate dehydrogenase kinase was found to possess a weak ATP hydrolytic activity, which required Glu-243 and His-239 similar to the kinase activity. Based on these observations, we propose a mechanism according to which the invariant glutamate residue (Glu-243) acts as a general base catalyst, which activates the hydroxyl group on a serine residue of the protein substrate for direct attack on the gamma phosphate. The glutamate residue in turn might be further polarized through interaction with the neighboring histidine residue (His-239).
Figures
References
-
- Randle PJ. Proc Nutr Soc. 1995;54:317–327. - PubMed
-
- Linn TC, Pettit FH, Reed LJ. Biochemistry. 1968;62:234–241.
-
- Stepp LR, Pettit FH, Yeaman SJ, Reed LJ. J Biol Chem. 1983;258:9454–9458. - PubMed
-
- Teague WM, Pettit FH, Wu TL, Silberman SR, Reed LJ. Biochemistry. 1982;21:5585–5592. - PubMed
-
- Popov KM, Kedishvili NY, Zhao Y, Gudi R, Harris RA. J Biol Chem. 1994;269:29720–29724. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
