

Two gamma2L subunit domains confer low Zn2+ sensitivity to ternary GABA(A) receptors
- PMID: 11283222
- PMCID: PMC2278531
- DOI: 10.1111/j.1469-7793.2001.0017g.x
Two gamma2L subunit domains confer low Zn2+ sensitivity to ternary GABA(A) receptors
Abstract
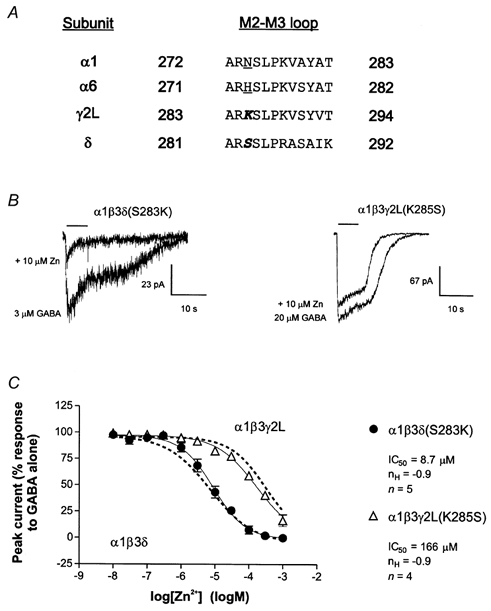
The sensitivity of GABAA receptors (GABARs) to Zn2+ inhibition depends on subunit composition. The predominant neuronal forms of mammalian GABARs, alpha(beta)gamma and, alpha(beta)delta are differentially sensitive to Zn2+ inhibition; alpha(beta)gamma receptors are substantially less sensitive than alpha(beta)delta receptors. Recently, functional domains involved in Zn2+ sensitivity have been identified in and subunits. Our aim in the present study was to localize functional domains of low Zn2+ sensitivity within gamma2L subunits. Chimeric subunits were constructed by progressively replacing the rat gamma2L subunit sequence with that of the rat delta subunit sequence. Whole-cell currents were recorded from mouse L929 fibroblasts coexpressing wild-type rat alpha1 and beta3 subunits with a chimeric delta-gamma2L subunit. Unlike alpha and beta subunits, the gamma2L subunit was found to contain a determinant of low Zn2+ sensitivity in the N-terminal extracellular region. In addition, we identified determinants in the M2 segment and the M2-M3 loop of the gamma2L subunit that are homologous to those found in beta and alpha subunits. We postulate that the interface between the latter two domains, which may form the outer vestibule of the channel, represents a single functional domain modulating Zn2+ sensitivity. Thus, the Zn2+ sensitivity of ternary GABARs appears to be determined by two functional domains, one in the N-terminal extracellular region and one near the outer mouth of the channel.
Figures







Similar articles
-
An N-terminal histidine is the primary determinant of alpha subunit-dependent Cu2+ sensitivity of alphabeta3gamma2L GABA(A) receptors.Mol Pharmacol. 2003 Nov;64(5):1145-52. doi: 10.1124/mol.64.5.1145. Mol Pharmacol. 2003. PMID: 14573764
-
The alpha 1 and alpha 6 subunit subtypes of the mammalian GABA(A) receptor confer distinct channel gating kinetics.J Physiol. 2004 Dec 1;561(Pt 2):433-48. doi: 10.1113/jphysiol.2003.051839. Epub 2004 Oct 7. J Physiol. 2004. PMID: 15579538 Free PMC article.
-
Zn2+ inhibition of recombinant GABAA receptors: an allosteric, state-dependent mechanism determined by the gamma-subunit.J Physiol. 1998 Feb 1;506 ( Pt 3)(Pt 3):609-25. doi: 10.1111/j.1469-7793.1998.609bv.x. J Physiol. 1998. PMID: 9503325 Free PMC article.
-
Potency of GABA at human recombinant GABA(A) receptors expressed in Xenopus oocytes: a mini review.Amino Acids. 2013 Apr;44(4):1139-49. doi: 10.1007/s00726-012-1456-y. Epub 2013 Feb 6. Amino Acids. 2013. PMID: 23385381 Review.
-
Multiple assembly signals in gamma-aminobutyric acid (type A) receptor subunits combine to drive receptor construction and composition.Biochem Soc Trans. 2003 Aug;31(Pt 4):875-9. doi: 10.1042/bst0310875. Biochem Soc Trans. 2003. PMID: 12887325 Review.
Cited by
-
Expression of human epileptic temporal lobe neurotransmitter receptors in Xenopus oocytes: An innovative approach to study epilepsy.Proc Natl Acad Sci U S A. 2002 Nov 12;99(23):15078-83. doi: 10.1073/pnas.232574499. Epub 2002 Oct 30. Proc Natl Acad Sci U S A. 2002. PMID: 12409614 Free PMC article.
-
Contrasting actions of a convulsant barbiturate and its anticonvulsant enantiomer on the α1 β3 γ2L GABAA receptor account for their in vivo effects.J Physiol. 2015 Nov 15;593(22):4943-61. doi: 10.1113/JP270971. J Physiol. 2015. PMID: 26378885 Free PMC article.
-
Electrophysiology of ionotropic GABA receptors.Cell Mol Life Sci. 2021 Jul;78(13):5341-5370. doi: 10.1007/s00018-021-03846-2. Epub 2021 Jun 1. Cell Mol Life Sci. 2021. PMID: 34061215 Free PMC article. Review.
-
Neurosteroid interactions with synaptic and extrasynaptic GABA(A) receptors: regulation of subunit plasticity, phasic and tonic inhibition, and neuronal network excitability.Psychopharmacology (Berl). 2013 Nov;230(2):151-88. doi: 10.1007/s00213-013-3276-5. Epub 2013 Sep 27. Psychopharmacology (Berl). 2013. PMID: 24071826 Free PMC article. Review.
-
γ-Aminobutyric acid type A (GABAA) receptor α subunits play a direct role in synaptic versus extrasynaptic targeting.J Biol Chem. 2012 Aug 10;287(33):27417-30. doi: 10.1074/jbc.M112.360461. Epub 2012 Jun 18. J Biol Chem. 2012. PMID: 22711532 Free PMC article.
References
-
- Bonnert TP, McKernan RM, Farrar S, le Bourdelle B, Heavens RP, Smith DW, Hewson L, Rigby MR, Sirinathsinghji DJ, Brown N, Wafford KA, Whiting PJ. Theta, a novel gamma-aminobutyric acid type A receptor subunit. Proceedings of the National Academy of Sciences of the USA. 1999;96:9891–9896. - PMC - PubMed
-
- Burgard EC, Tietz EI, Neelands TR, Macdonald RL. Properties of recombinant γ-aminobutyric acidA receptor isoforms containing the α5 subunit subtype. Molecular Pharmacology. 1996;50:119–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources