

FtsZ ring formation at the chloroplast division site in plants
- PMID: 11285278
- PMCID: PMC2185535
- DOI: 10.1083/jcb.153.1.111
FtsZ ring formation at the chloroplast division site in plants
Abstract
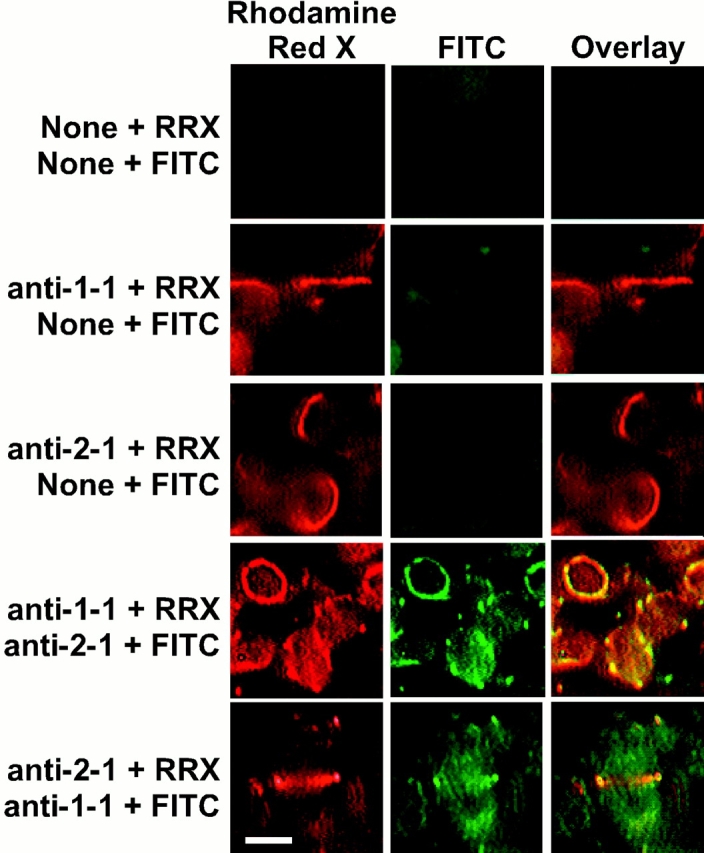
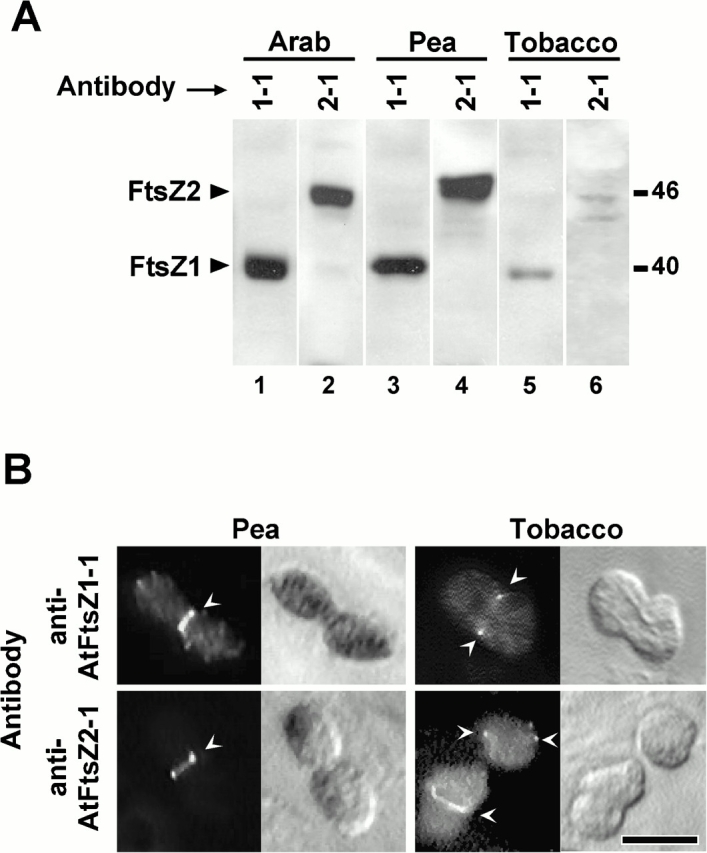
Among the events that accompanied the evolution of chloroplasts from their endosymbiotic ancestors was the host cell recruitment of the prokaryotic cell division protein FtsZ to function in chloroplast division. FtsZ, a structural homologue of tubulin, mediates cell division in bacteria by assembling into a ring at the midcell division site. In higher plants, two nuclear-encoded forms of FtsZ, FtsZ1 and FtsZ2, play essential and functionally distinct roles in chloroplast division, but whether this involves ring formation at the division site has not been determined previously. Using immunofluorescence microscopy and expression of green fluorescent protein fusion proteins in Arabidopsis thaliana, we demonstrate here that FtsZ1 and FtsZ2 localize to coaligned rings at the chloroplast midpoint. Antibodies specific for recognition of FtsZ1 or FtsZ2 proteins in Arabidopsis also recognize related polypeptides and detect midplastid rings in pea and tobacco, suggesting that midplastid ring formation by FtsZ1 and FtsZ2 is universal among flowering plants. Perturbation in the level of either protein in transgenic plants is accompanied by plastid division defects and assembly of FtsZ1 and FtsZ2 into filaments and filament networks not observed in wild-type, suggesting that previously described FtsZ-containing cytoskeletal-like networks in chloroplasts may be artifacts of FtsZ overexpression.
Figures





References
-
- Beech P.L., Gilson P.R. FtsZ and organelle division in protists. Protist. 2000;151:11–16. - PubMed
-
- Beech P.L., Nheu T., Schultz T., Herbert S., Lithgow T., Gilson P.R., McFadden G.I. Mitochondrial FtsZ in a chromophyte alga. Science. 2000;287:1276–1279. - PubMed
-
- Bi E., Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli . Nature. 1991;354:161–164. - PubMed
-
- Bramhill D. Bacterial cell division. Annu. Rev. Cell Dev. Biol. 1997;13:395–424. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
