

Role of NF-Y in in vivo regulation of the gamma-globin gene
- PMID: 11287613
- PMCID: PMC86936
- DOI: 10.1128/MCB.21.9.3083-3095.2001
Role of NF-Y in in vivo regulation of the gamma-globin gene
Abstract
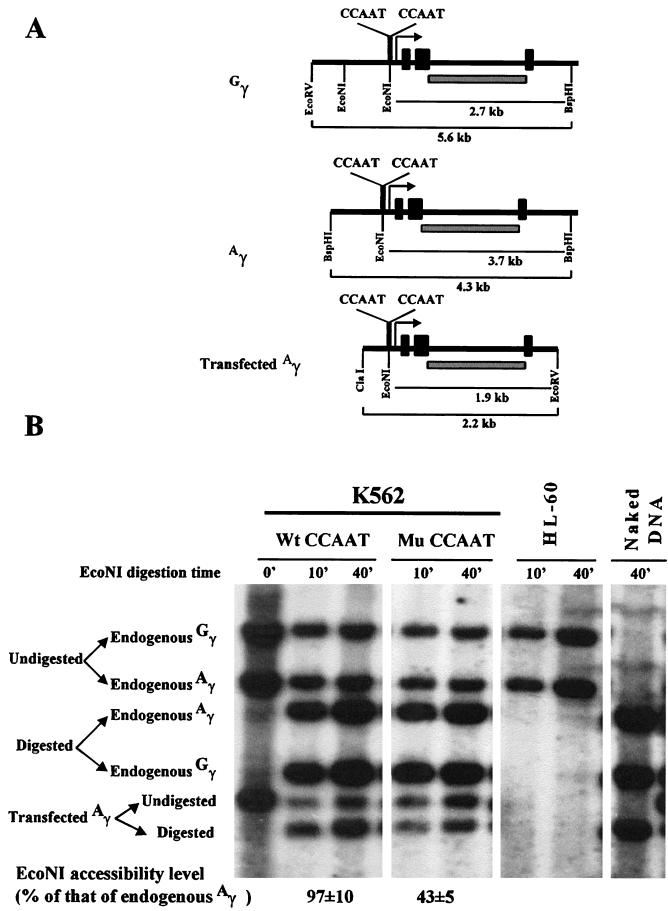
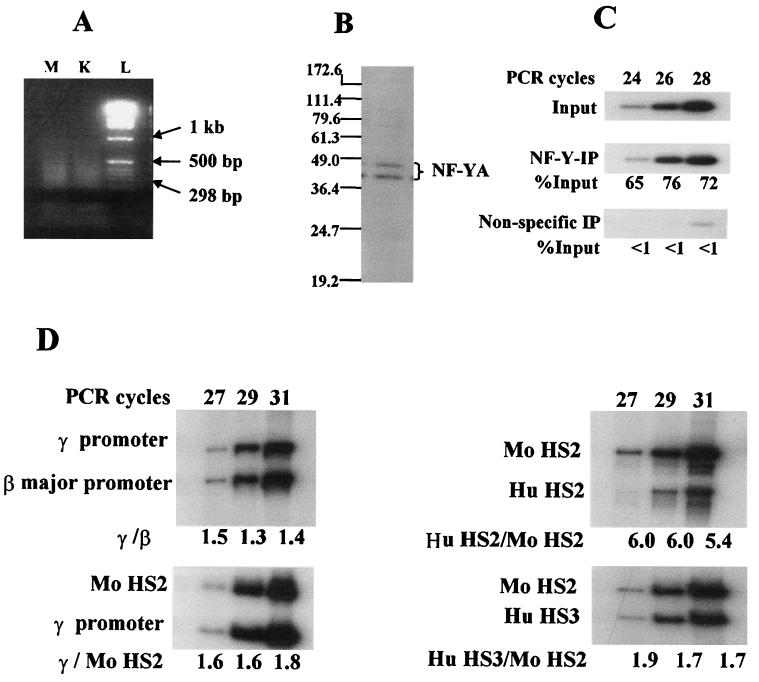
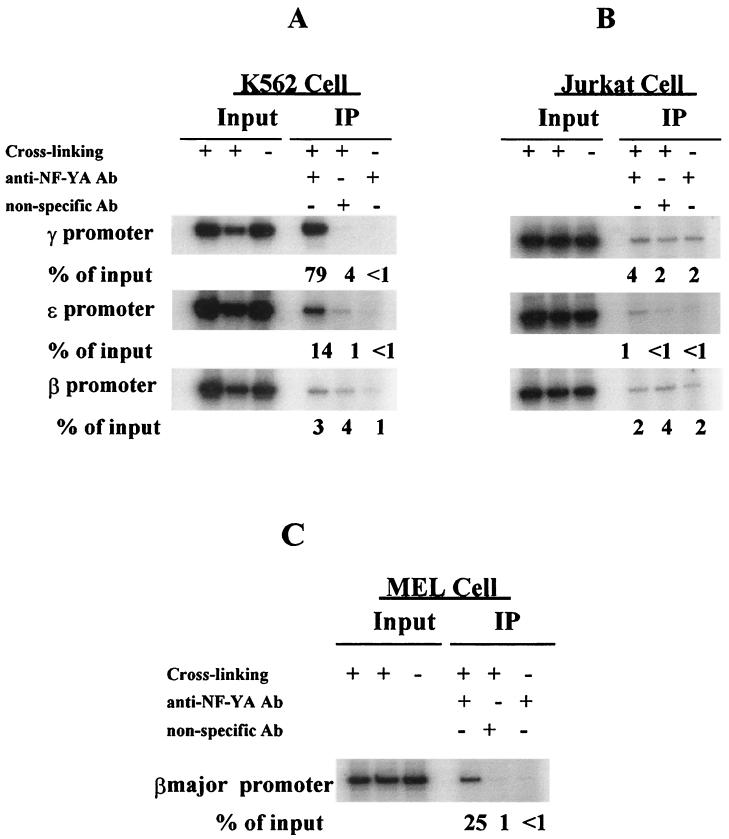
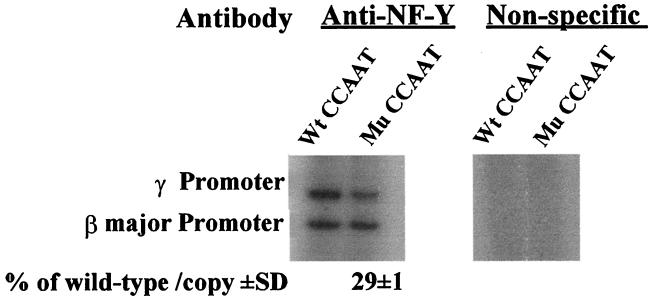
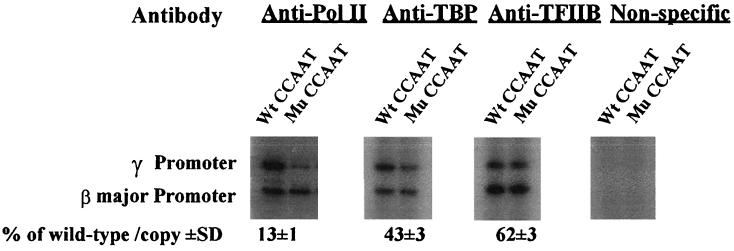
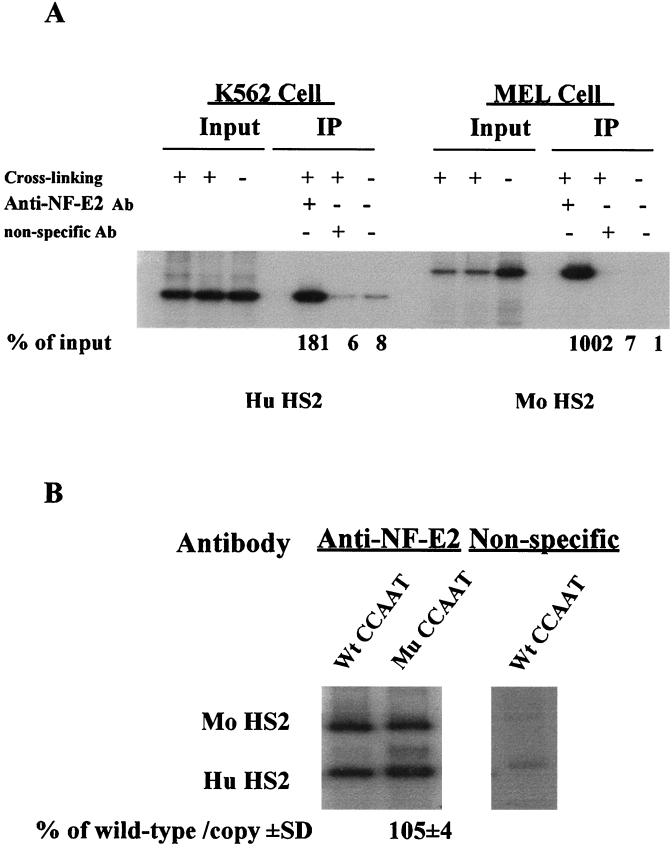
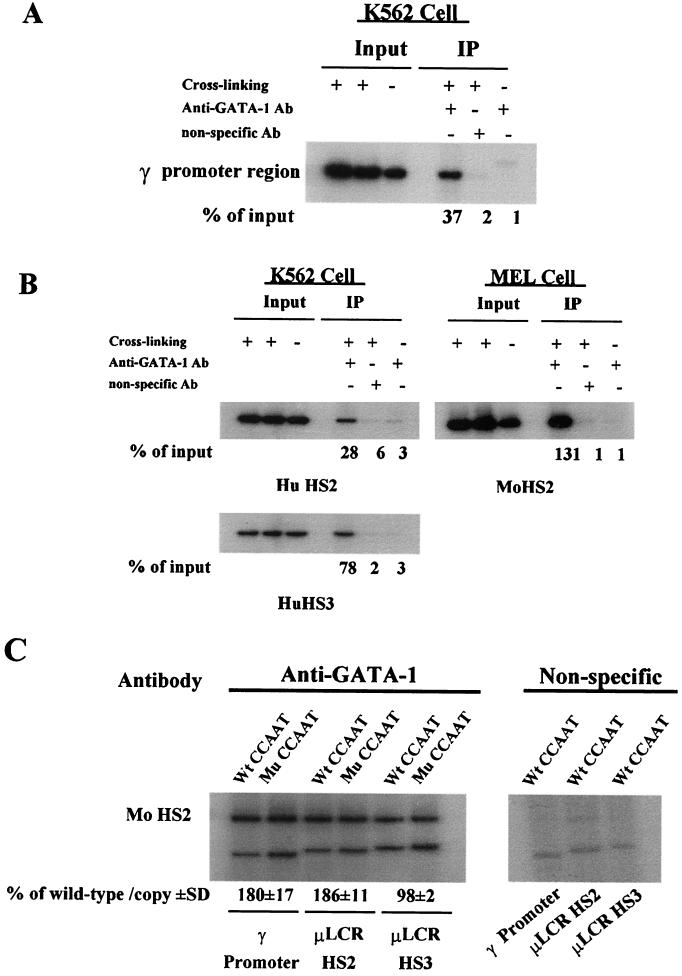
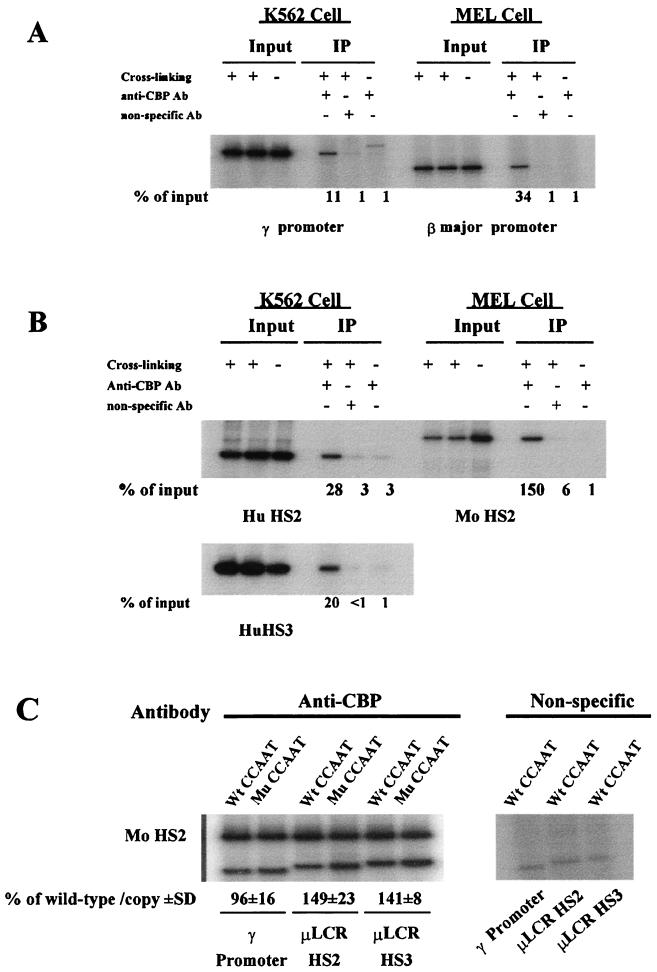
The duplicated CCAAT box is required for gamma gene expression. We report here that the transcriptional factor NF-Y is recruited to the duplicated CCAAT box in vivo. A mutation of the duplicated CCAAT box that severely disrupts the NF-Y binding also reduces the accessibility level of the gamma gene promoter, affects the assembly of basal transcriptional machinery, and increases the recruitment of GATA-1 to the locus control region (LCR) and the proximal promoter and the recruitment of transcription cofactor CBP/p300 to the LCR. These findings suggest that recruitment of NF-Y to the duplicated CCAAT box plays a role in the chromatin opening of the gamma gene promoter as well as in the communication between the gamma gene promoter and the LCR.
Figures
References
-
- Armstrong J A, Bieker J J, Emerson B M. A SWI/SNF-related chromatin remodeling complex, E-RC1, is required for tissue-specific transcriptional regulation by EKLF in vitro. Cell. 1998;95:93–104. - PubMed
-
- Berry M, Grosveld F, Dillon N. A single point mutation is the cause of the Greek form of hereditary persistence of fetal haemoglobin. Nature. 1992;358:499–502. - PubMed
-
- Blobel G A. CREB-binding protein and p300: molecular integrators of hematopoietic transcription. Blood. 2000;95:745–755. - PubMed
-
- Bulger M, Goudine M. Looping versus linking: toward a model for long-distance gene activation. Genes Dev. 1999;13:2465–2477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous