

RAC1 regulates adherens junctions through endocytosis of E-cadherin
- PMID: 11294891
- PMCID: PMC32271
- DOI: 10.1091/mbc.12.4.847
RAC1 regulates adherens junctions through endocytosis of E-cadherin
Abstract
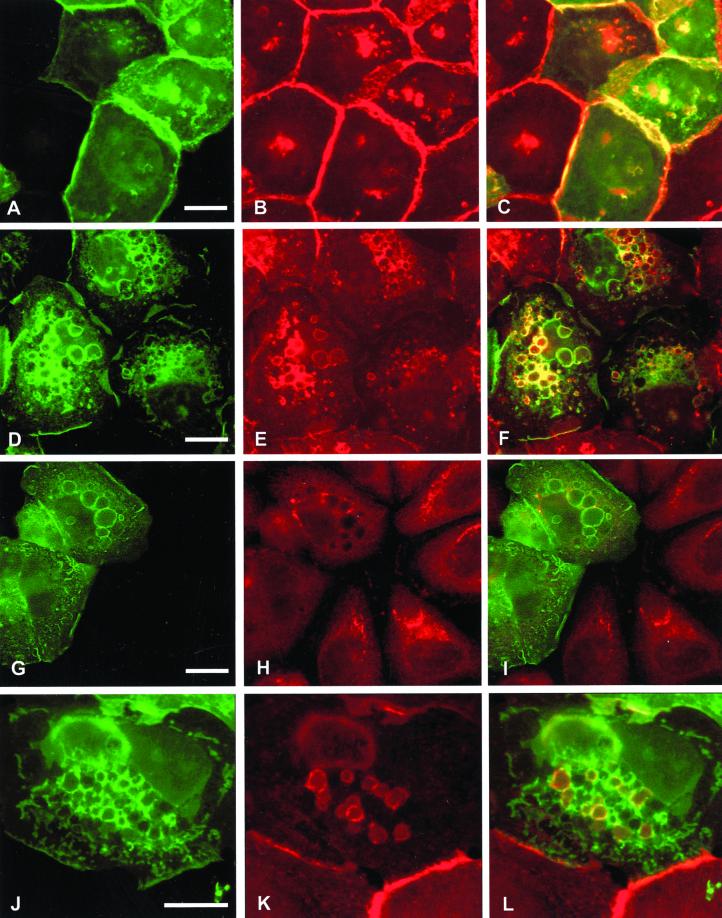
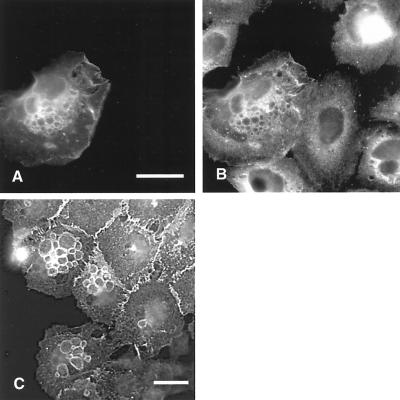
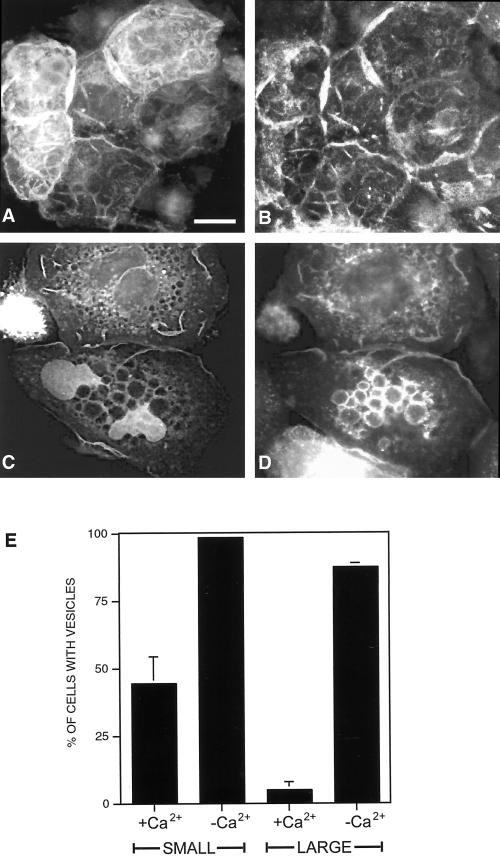
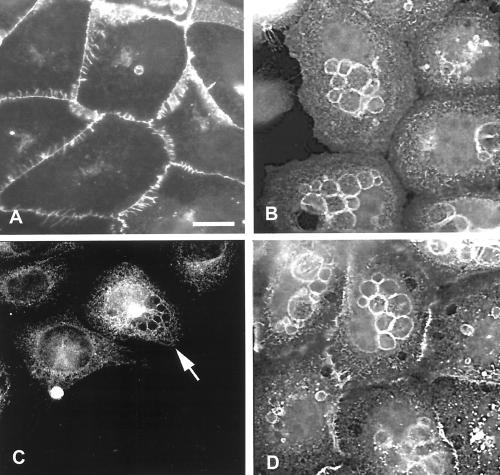
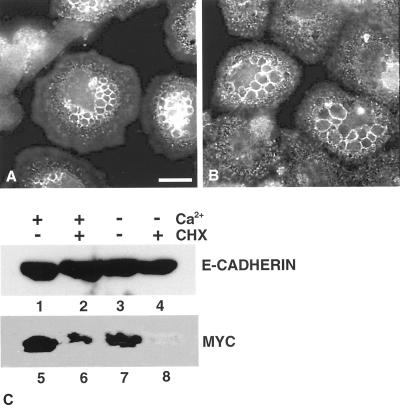
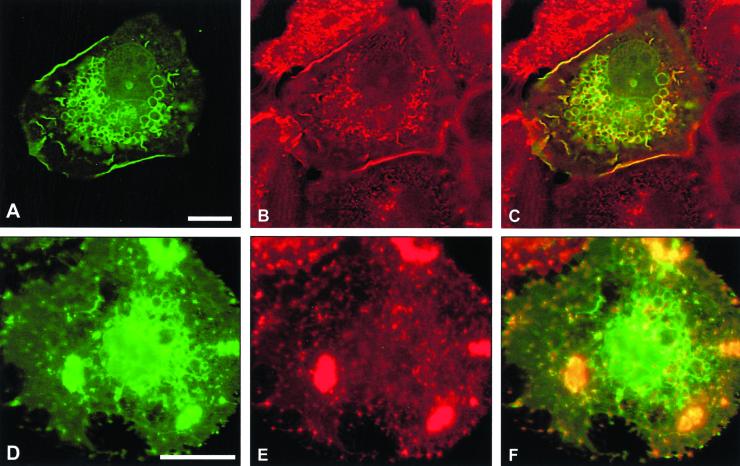
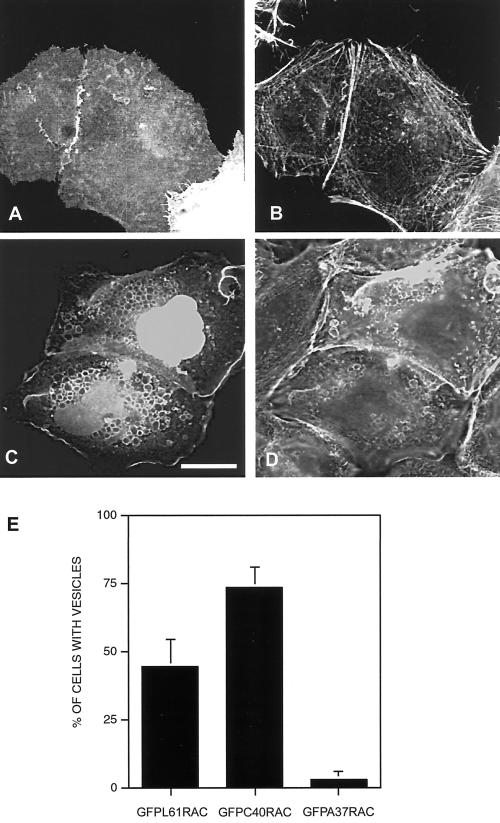
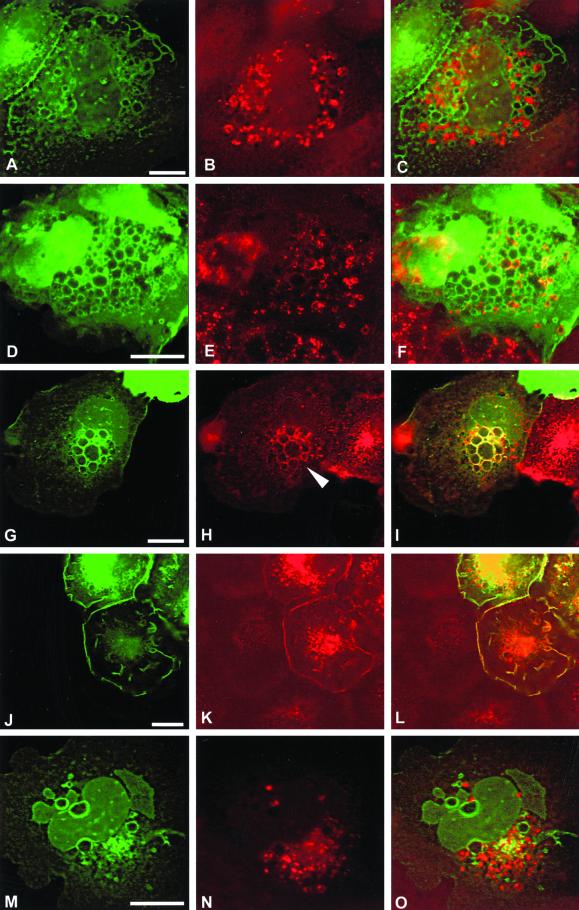
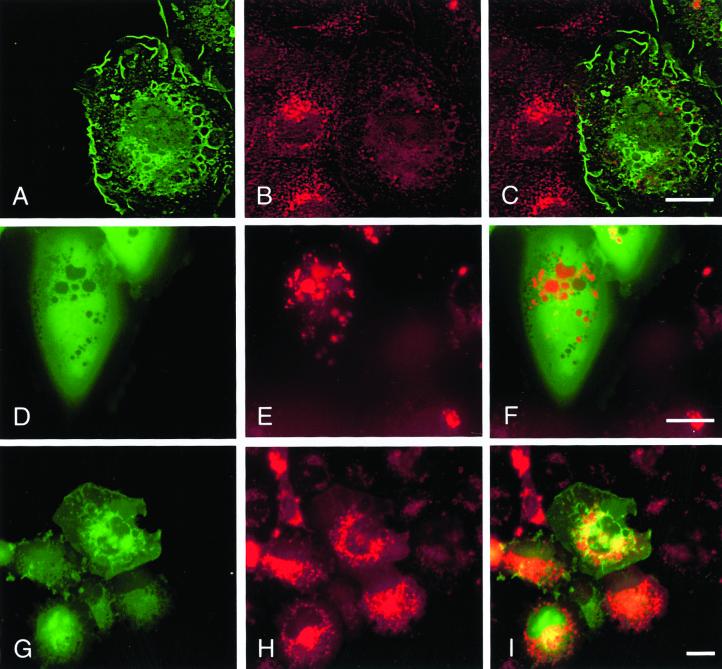
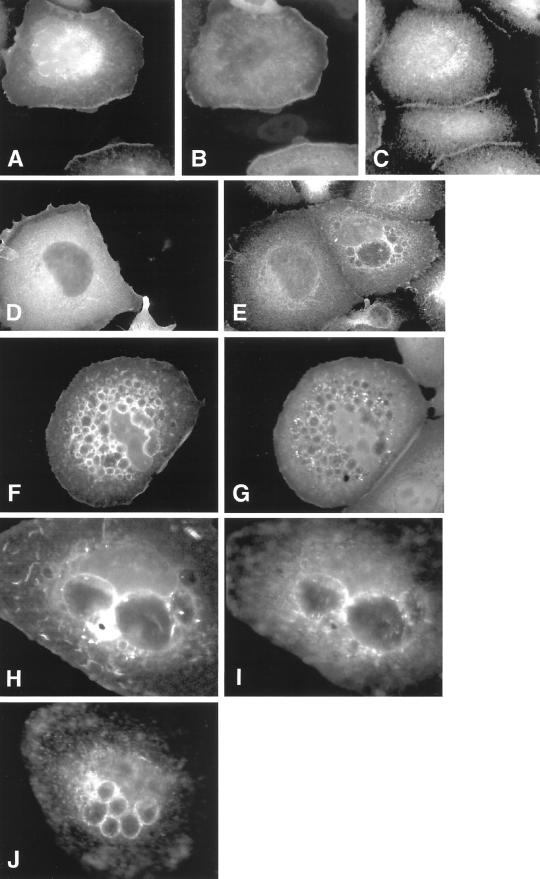
The establishment of cadherin-dependent cell-cell contacts in human epidermal keratinocytes are known to be regulated by the Rac1 small GTP-binding protein, although the mechanisms by which Rac1 participates in the assembly or disruption of cell-cell adhesion are not well understood. In this study we utilized green fluorescent protein (GFP)-tagged Rac1 expression vectors to examine the subcellular distribution of Rac1 and its effects on E-cadherin-mediated cell-cell adhesion. Microinjection of keratinocytes with constitutively active Rac1 resulted in cell spreading and disruption of cell-cell contacts. The ability of Rac1 to disrupt cell-cell adhesion was dependent on colony size, with large established colonies being resistant to the effects of active Rac1. Disruption of cell-cell contacts in small preconfluent colonies was achieved through the selective recruitment of E-cadherin-catenin complexes to the perimeter of multiple large intracellular vesicles, which were bounded by GFP-tagged L61Rac1. Similar vesicles were observed in noninjected keratinocytes when cell-cell adhesion was disrupted by removal of extracellular calcium or with the use of an E-cadherin blocking antibody. Moreover, formation of these structures in noninjected keratinocytes was dependent on endogenous Rac1 activity. Expression of GFP-tagged effector mutants of Rac1 in keratinocytes demonstrated that reorganization of the actin cytoskeleton was important for vesicle formation. Characterization of these Rac1-induced vesicles revealed that they were endosomal in nature and tightly colocalized with the transferrin receptor, a marker for recycling endosomes. Expression of GFP-L61Rac1 inhibited uptake of transferrin-biotin, suggesting that the endocytosis of E-cadherin was a clathrin-independent mechanism. This was supported by the observation that caveolin, but not clathrin, localized around these structures. Furthermore, an inhibitory form of dynamin, known to inhibit internalization of caveolae, inhibited formation of cadherin vesicles. Our data suggest that Rac1 regulates adherens junctions via clathrin independent endocytosis of E-cadherin.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
