

Depletion of acyl-coenzyme A-binding protein affects sphingolipid synthesis and causes vesicle accumulation and membrane defects in Saccharomyces cerevisiae
- PMID: 11294913
- PMCID: PMC32293
- DOI: 10.1091/mbc.12.4.1147
Depletion of acyl-coenzyme A-binding protein affects sphingolipid synthesis and causes vesicle accumulation and membrane defects in Saccharomyces cerevisiae
Abstract
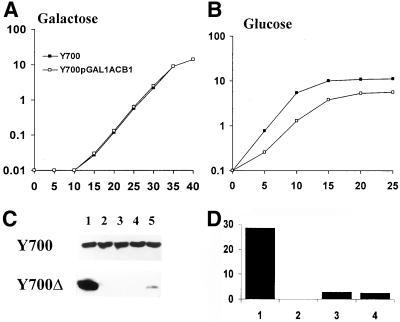
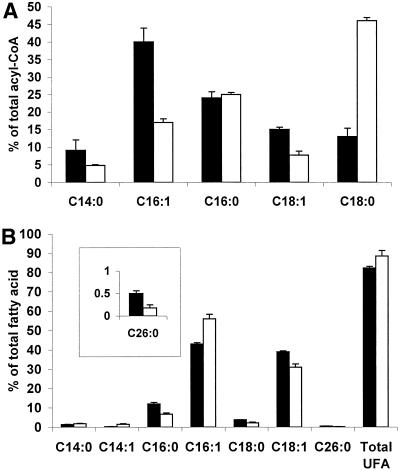
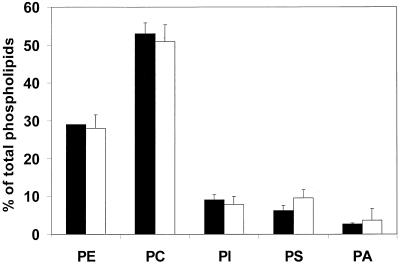
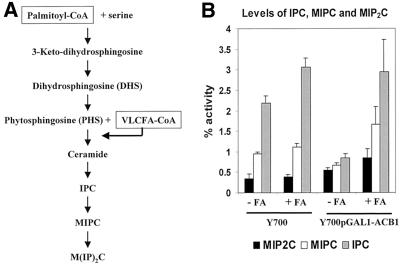
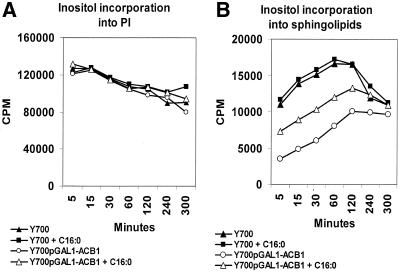
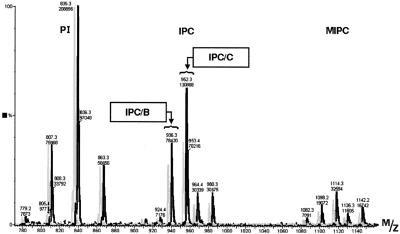
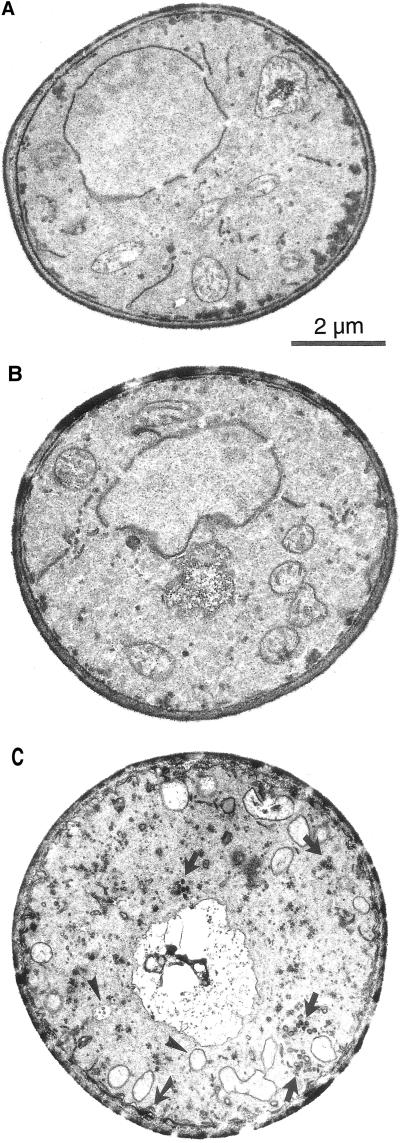
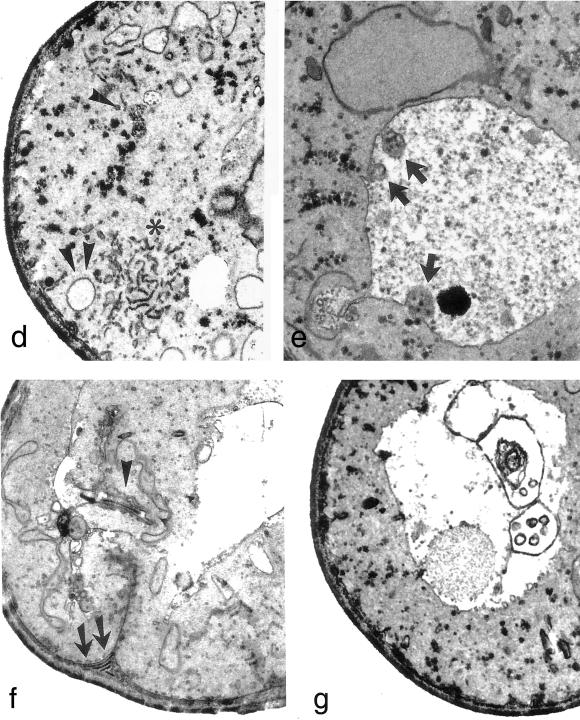
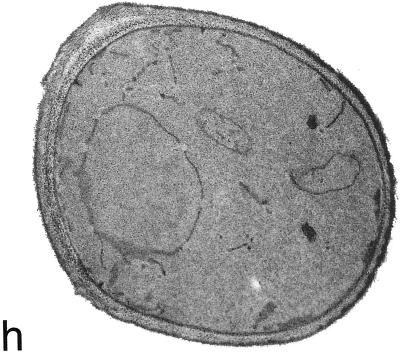
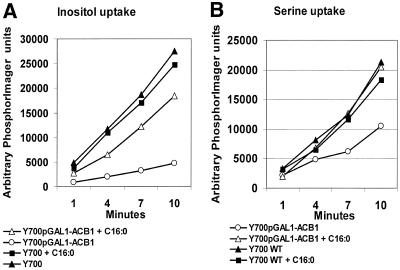
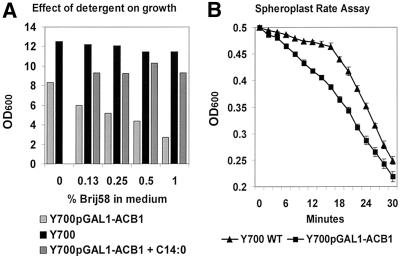
Deletion of the yeast gene ACB1 encoding Acb1p, the yeast homologue of the acyl-CoA-binding protein (ACBP), resulted in a slower growing phenotype that adapted into a faster growing phenotype with a frequency >1:10(5). A conditional knockout strain (Y700pGAL1-ACB1) with the ACB1 gene under control of the GAL1 promoter exhibited an altered acyl-CoA profile with a threefold increase in the relative content of C18:0-CoA, without affecting total acyl-CoA level as previously reported for an adapted acb1Delta strain. Depletion of Acb1p did not affect the general phospholipid pattern, the rate of phospholipid synthesis, or the turnover of individual phospholipid classes, indicating that Acb1p is not required for general glycerolipid synthesis. In contrast, cells depleted for Acb1p showed a dramatically reduced content of C26:0 in total fatty acids and the sphingolipid synthesis was reduced by 50-70%. The reduced incorporation of [(3)H]myo-inositol into sphingolipids was due to a reduced incorporation into inositol-phosphoceramide and mannose-inositol-phosphoceramide only, a pattern that is characteristic for cells with aberrant endoplasmic reticulum to Golgi transport. The plasma membrane of the Acb1p-depleted strain contained increased levels of inositol-phosphoceramide and mannose-inositol-phosphoceramide and lysophospholipids. Acb1p-depleted cells accumulated 50- to 60-nm vesicles and autophagocytotic like bodies and showed strongly perturbed plasma membrane structures. The present results strongly suggest that Acb1p plays an important role in fatty acid elongation and membrane assembly and organization.
Figures

References
-
- Borsting C, Hummel R, Schultz ER, Rose TM, Pedersen MB, Knudsen J, Kristiansen K. Saccharomyces carlsbergensis contains two functional genes encoding the acyl-CoA binding protein, one similar to the ACB1 gene from S. cerevisiae and one identical to the ACB1 gene from S. monacensis. Yeast. 1997;13:1409–1421. - PubMed
-
- Choi JY, Stukey J, Hwang SY, Martin CE. Regulatory elements that control transcription activation and unsaturated fatty acid-mediated repression of the Saccharomyces cerevisiae OLE1 gene. J Biol Chem. 1996;271:3581–3589. - PubMed
-
- Færgeman NJ, Sigurskjold BW, Kragelund BB, Andersen KV, Knudsen J. Thermodynamics of ligand binding to acyl-coenzyme A binding protein studied by titration calorimetry. Biochemistry. 1996;35:14118–14126. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
