

mAKAP assembles a protein kinase A/PDE4 phosphodiesterase cAMP signaling module
- PMID: 11296225
- PMCID: PMC125429
- DOI: 10.1093/emboj/20.8.1921
mAKAP assembles a protein kinase A/PDE4 phosphodiesterase cAMP signaling module
Abstract
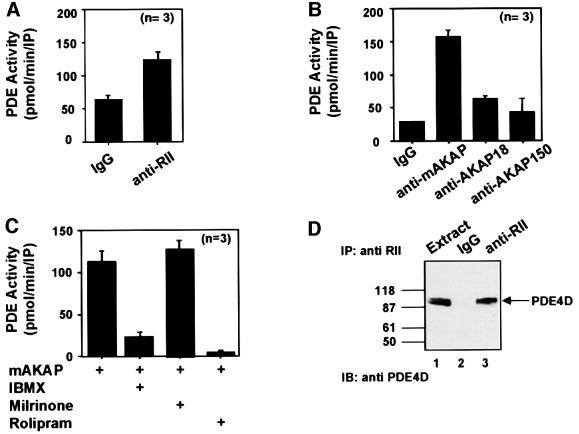
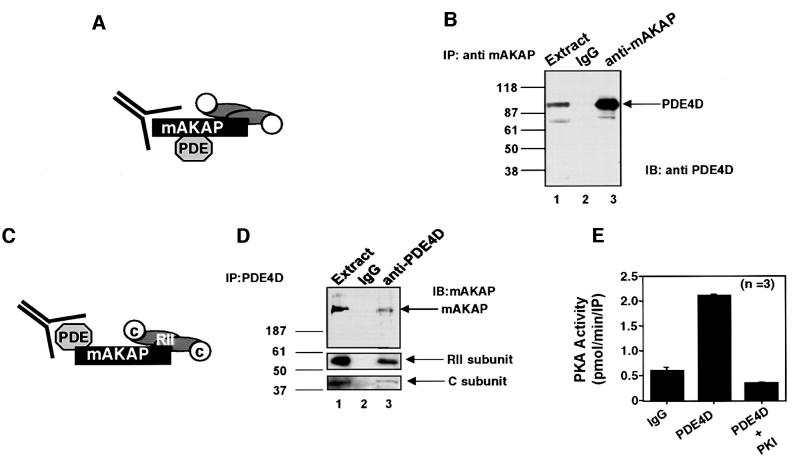
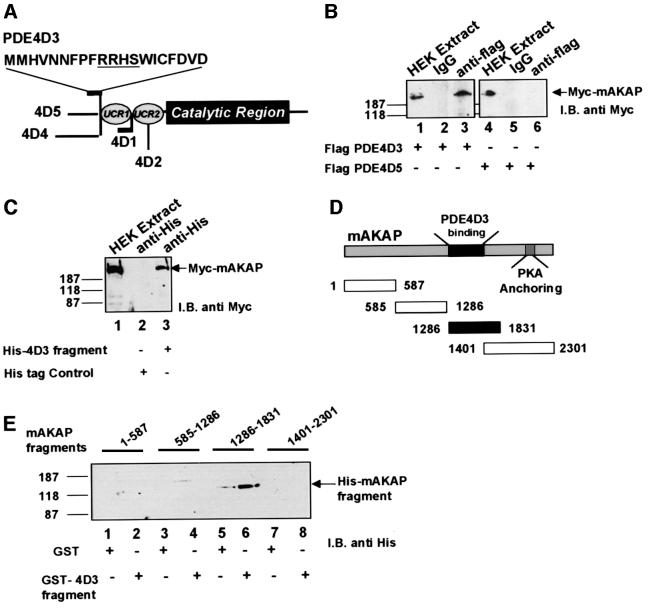
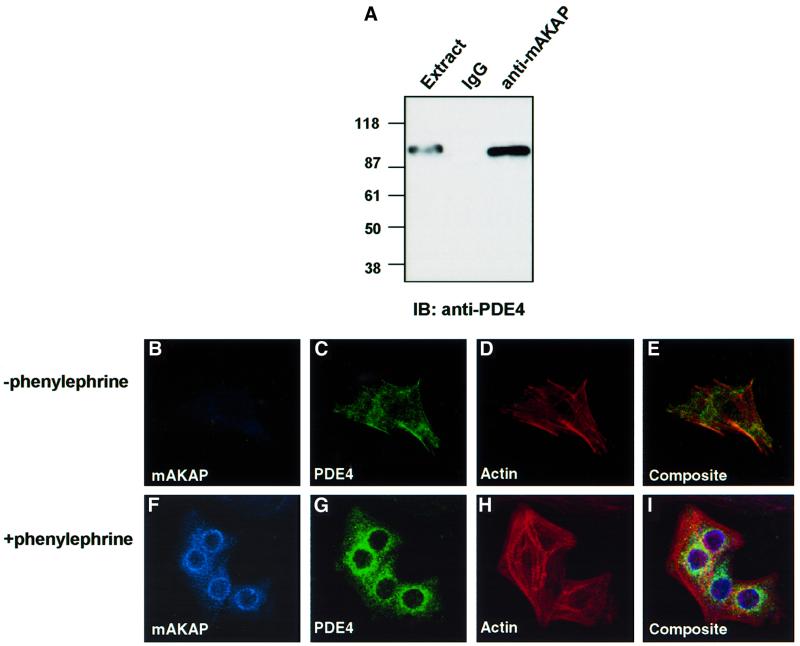
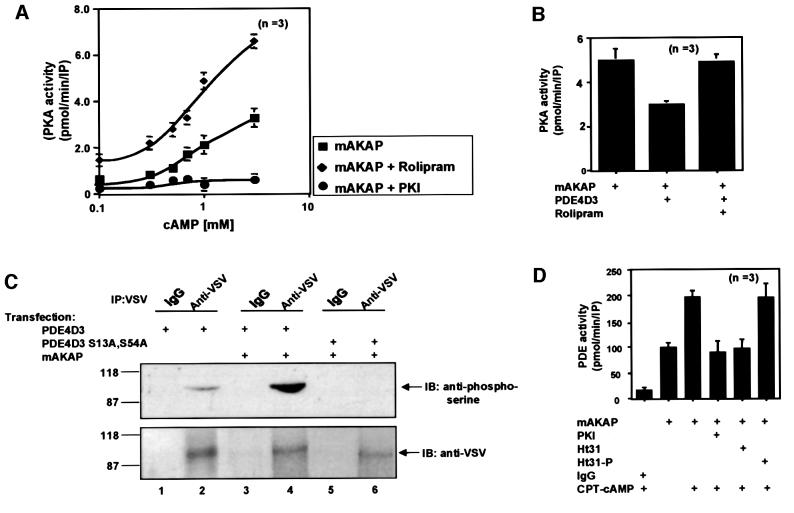
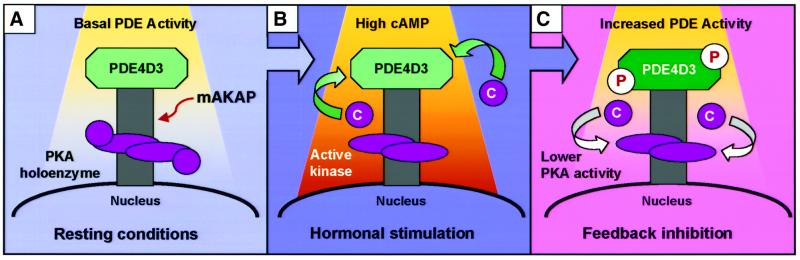
Spatiotemporal regulation of protein kinase A (PKA) activity involves the manipulation of compartmentalized cAMP pools. Now we demonstrate that the muscle-selective A-kinase anchoring protein, mAKAP, maintains a cAMP signaling module, including PKA and the rolipram-inhibited cAMP-specific phosphodiesterase (PDE4D3) in heart tissues. Functional analyses indicate that tonic PDE4D3 activity reduces the activity of the anchored PKA holoenzyme, whereas kinase activation stimulates mAKAP-associated phosphodiesterase activity. Disruption of PKA- mAKAP interaction prevents this enhancement of PDE4D3 activity, suggesting that the proximity of both enzymes in the mAKAP signaling complex forms a negative feedback loop to restore basal cAMP levels.
Figures
References
-
- Beard M.B., O’Connell,J.C., Bolger,G.B. and Houslay,M.D. (1999) The unique N-terminal domain of the cAMP phosphodiesterase PDE4D4 allows for interaction with specific SH3 domains. FEBS Lett., 460, 173–177. - PubMed
-
- Beard M.B., Olsen,A.E., Jones,R.E., Erdogan,S., Houslay,M.D. and Bolger,G.B. (2000) UCR1 and UCR2 domains unique to the cAMP-specific phosphodiesterase family form a discrete module via electrostatic interactions. J. Biol. Chem., 275, 10349–10358. - PubMed
-
- Beavo J.A. (1995) Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiol. Rev., 75, 725–748. - PubMed
-
- Beavo J.A., Bechtel,P.J. and Krebs,E.G. (1974) Preparation of homogenous cyclic AMP-dependent protein kinase(s) and its subunits from rabbit skeletal muscle. Methods Enzymol., 38, 299–308. - PubMed
-
- Beavo J.A., Conti,M. and Heaslip,R.J. (1994) Multiple cyclic nucleotide phosphodiesterases. Mol. Pharmacol., 46, 399–405. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
