

Saccharomyces cerevisiae Elongator mutations confer resistance to the Kluyveromyces lactis zymocin
- PMID: 11296232
- PMCID: PMC125238
- DOI: 10.1093/emboj/20.8.1993
Saccharomyces cerevisiae Elongator mutations confer resistance to the Kluyveromyces lactis zymocin
Abstract
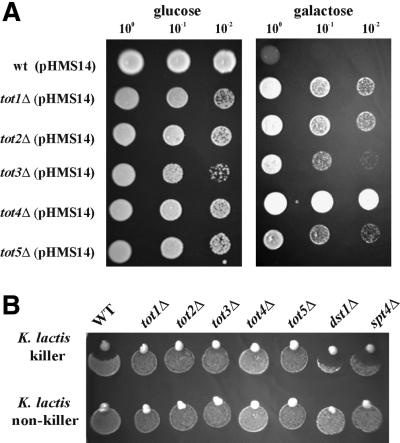
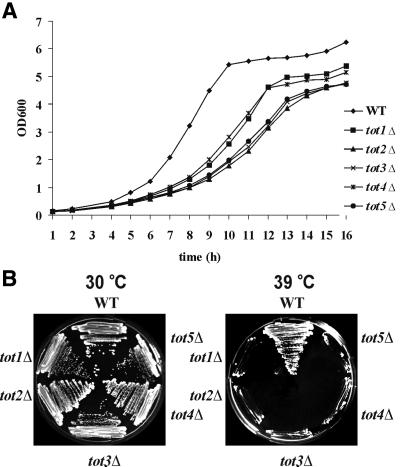
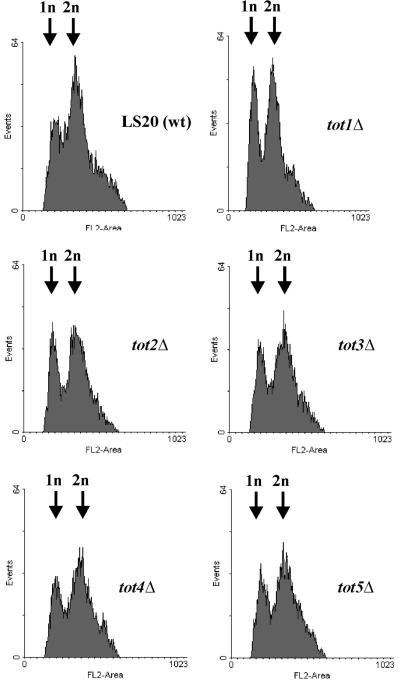
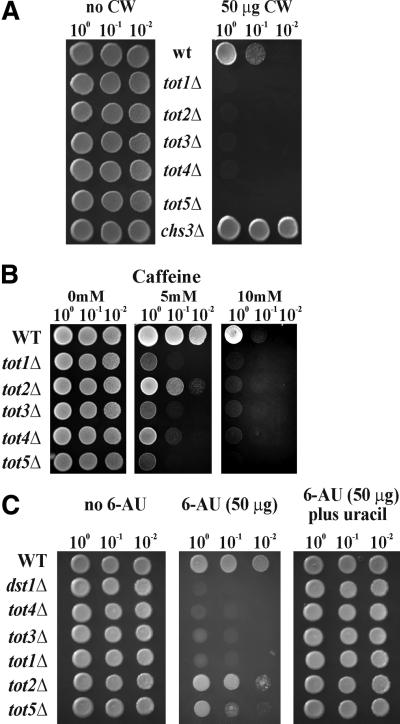
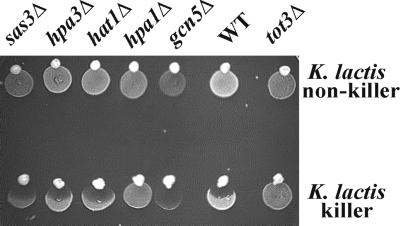
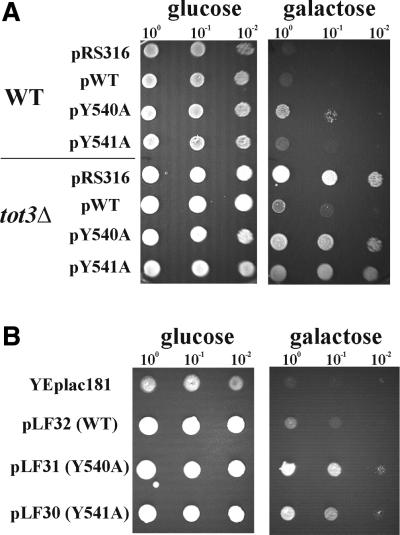
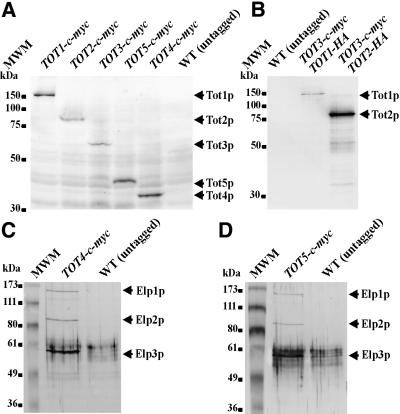
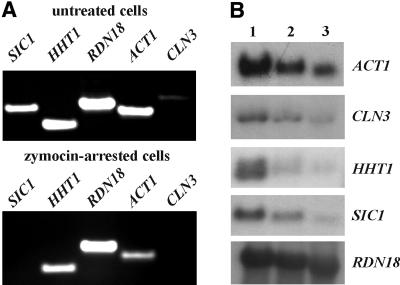
Kluyveromyces lactis killer strains secrete a zymocin complex that inhibits proliferation of sensitive yeast genera including Saccharomyces cerevisiae. In search of the putative toxin target (TOT), we used mTn3:: tagging to isolate zymocin-resistant tot mutants from budding yeast. Of these we identified the TOT1, TOT2 and TOT3 genes (isoallelic with ELP1, ELP2 and ELP3, respectively) coding for the histone acetyltransferase (HAT)-associated Elongator complex of RNA polymerase II holoenzyme. Other than the typical elp ts-phenotype, tot phenocopies hypersensitivity towards caffeine and Calcofluor White as well as slow growth and a G(1) cell cycle delay. In addition, TOT4 and TOT5 (isoallelic with KTI12 and IKI1, respectively) code for components that associate with ELONGATOR: Intriguingly, strains lacking non-Elongator HATs (gcn5, hat1, hpa3 and sas3) or non-Elongator transcription elongation factors TFIIS (dst1) and Spt4p (spt4) cannot confer resistance towards the K.lactis zymocin, thus providing evidence that Elongator equals TOT and that Elongator plays an important role in signalling toxicity of the K.lactis zymocin.
Figures
References
-
- Ahmed A., Sesti,F., Ilan,N., Shih,T.M., Sturley,S.L. and Goldstein,S.A. (1999) A molecular target for viral killer toxin: TOK1 potassium channels. Cell, 99, 283–291. - PubMed
-
- Becker D.M., and Guarente,L. (1991) High-efficiency transformation of yeast by electroporation. Methods Enzymol., 194, 182–187. - PubMed
-
- Berben G., Dumont,J., Gilliquet,V., Bolle,P.A. and Hilger,F. (1991) The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast, 7, 475–477. - PubMed
-
- Brown C.E., Lechner,T., Howe,L. and Workman,J.L. (2000) The many HATs of transcription coactivators. Trends Biochem. Sci., 25, 15–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
