

MOT1-catalyzed TBP-DNA disruption: uncoupling DNA conformational change and role of upstream DNA
- PMID: 11296235
- PMCID: PMC125425
- DOI: 10.1093/emboj/20.8.2028
MOT1-catalyzed TBP-DNA disruption: uncoupling DNA conformational change and role of upstream DNA
Abstract
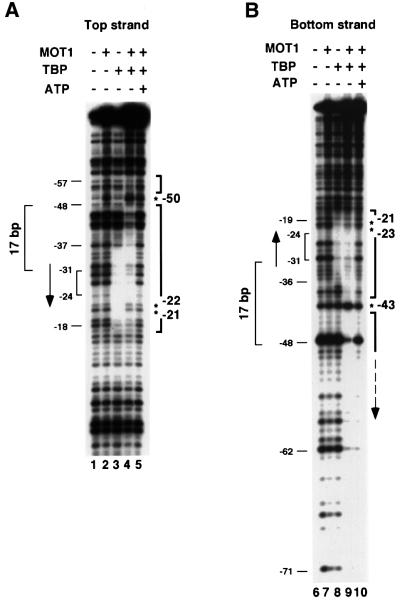
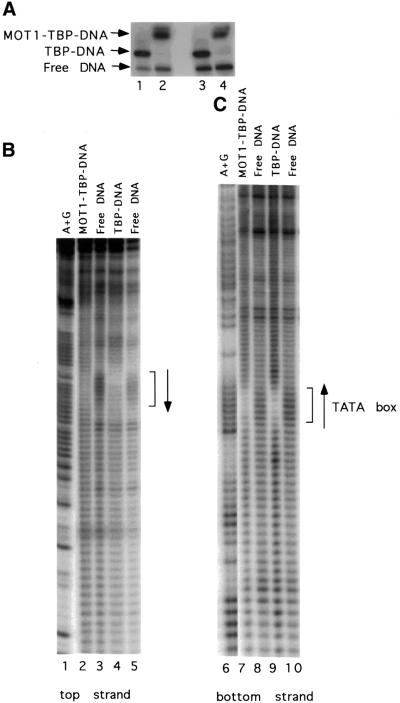
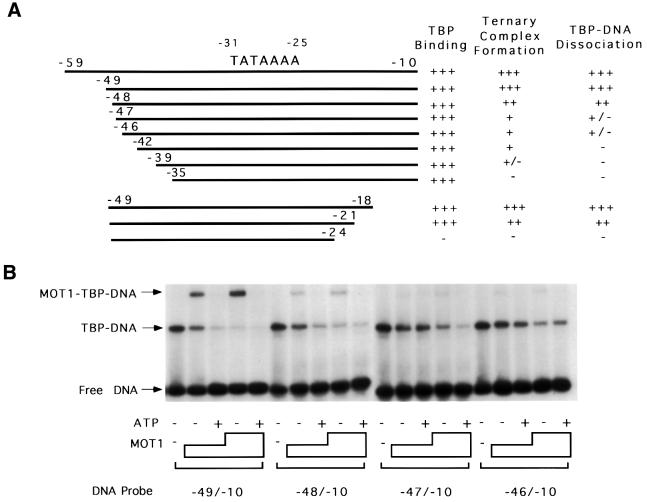
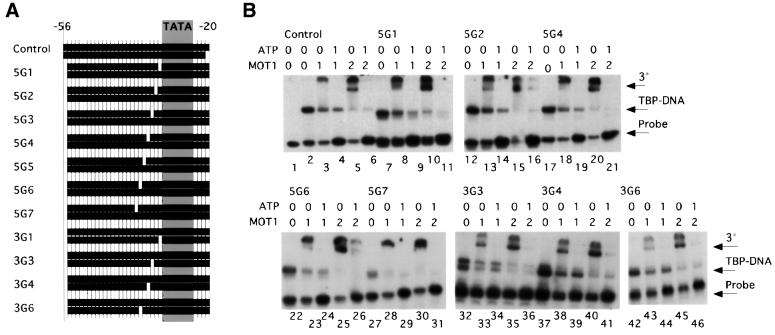
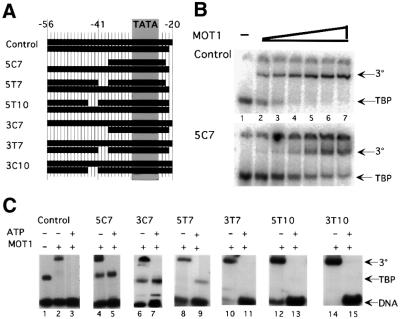
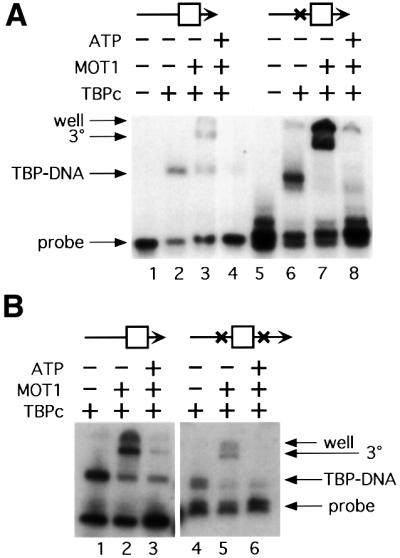
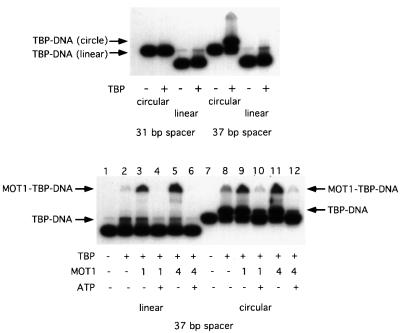
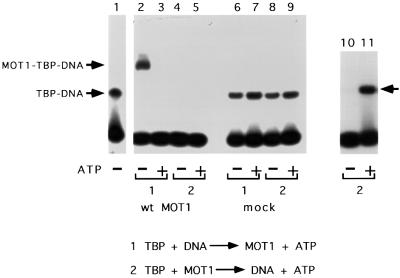
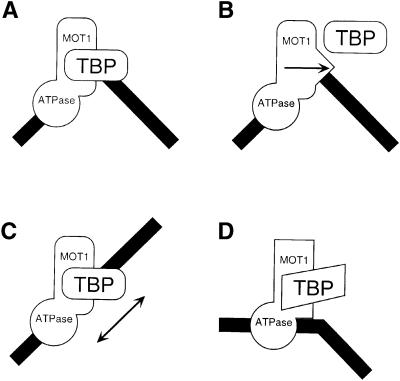
SNF2/SWI2-related ATPases employ ATP hydrolysis to disrupt protein-DNA interactions, but how ATP hydrolysis is coupled to disruption is not understood. Here we examine the mechanism of action of MOT1, a yeast SNF2/SWI2-related ATPase that uses ATP hydrolysis to remove TATA binding protein (TBP) from DNA. MOT1 function requires a 17 bp DNA 'handle' upstream of the TATA box, which must be double stranded. Remarkably, MOT1-catalyzed disruption of TBP-DNA does not appear to require DNA strand separation, DNA bending or twisting of the DNA helix. Thus, TBP-DNA disruption is accomplished in a reaction apparently not driven by a change in DNA structure. MOT1 action is supported by DNA templates in which the handle is connected to the TATA box via single-stranded DNA, indicating that the upstream duplex DNA can be conformationally uncoupled from the TATA box. Combining these results with proposed similarities between SNF2/SWI2 ATPases and helicases, we suggest that MOT1 uses ATP hydrolysis to translocate along the handle and thereby disrupt interactions between TBP and DNA.
Figures
Similar articles
-
Molecular analysis of the SNF2/SWI2 protein family member MOT1, an ATP-driven enzyme that dissociates TATA-binding protein from DNA.Mol Cell Biol. 1997 Aug;17(8):4842-51. doi: 10.1128/MCB.17.8.4842. Mol Cell Biol. 1997. PMID: 9234740 Free PMC article.
-
Testing for DNA tracking by MOT1, a SNF2/SWI2 protein family member.Mol Cell Biol. 1999 Jan;19(1):412-23. doi: 10.1128/MCB.19.1.412. Mol Cell Biol. 1999. PMID: 9858565 Free PMC article.
-
Snf2/Swi2-related ATPase Mot1 drives displacement of TATA-binding protein by gripping DNA.EMBO J. 2006 Apr 5;25(7):1492-504. doi: 10.1038/sj.emboj.7601050. Epub 2006 Mar 16. EMBO J. 2006. PMID: 16541100 Free PMC article.
-
One small step for Mot1; one giant leap for other Swi2/Snf2 enzymes?Biochim Biophys Acta. 2011 Sep;1809(9):488-96. doi: 10.1016/j.bbagrm.2011.05.012. Epub 2011 May 30. Biochim Biophys Acta. 2011. PMID: 21658482 Free PMC article. Review.
-
X-ray crystallographic studies of eukaryotic transcription initiation factors.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):483-9. doi: 10.1098/rstb.1996.0046. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735270 Review.
Cited by
-
Structural basis for recognition and remodeling of the TBP:DNA:NC2 complex by Mot1.Elife. 2015 Aug 10;4:e07432. doi: 10.7554/eLife.07432. Elife. 2015. PMID: 26258880 Free PMC article.
-
Crystal structure of the full Swi2/Snf2 remodeler Mot1 in the resting state.Elife. 2018 Oct 5;7:e37774. doi: 10.7554/eLife.37774. Elife. 2018. PMID: 30289385 Free PMC article.
-
Structural basis for TBP displacement from TATA box DNA by the Swi2/Snf2 ATPase Mot1.Nat Struct Mol Biol. 2023 May;30(5):640-649. doi: 10.1038/s41594-023-00966-0. Epub 2023 Apr 27. Nat Struct Mol Biol. 2023. PMID: 37106137 Free PMC article.
-
Function and structural organization of Mot1 bound to a natural target promoter.J Biol Chem. 2008 Sep 5;283(36):24935-48. doi: 10.1074/jbc.M803749200. Epub 2008 Jul 7. J Biol Chem. 2008. PMID: 18606810 Free PMC article.
-
Mot1 associates with transcriptionally active promoters and inhibits association of NC2 in Saccharomyces cerevisiae.Mol Cell Biol. 2002 Dec;22(23):8122-34. doi: 10.1128/MCB.22.23.8122-8134.2002. Mol Cell Biol. 2002. PMID: 12417716 Free PMC article.
References
-
- Adamkewicz J.I., Mueller,C.G.F., Hansen,K.E., Prud’homme,W.A. and Thorner,J. (2000) Purification and enzymic properties of Mot1 ATPase, a regulator of basal transcription in the yeast Saccharomyces cerevisiae. J. Biol. Chem., 275, 21158–21168. - PubMed
-
- Auble D.T. and Hahn,S. (1993) An ATP-dependent inhibitor of TBP binding to DNA. Genes Dev., 7, 844–856. - PubMed
-
- Auble D.T., Hansen,K.E., Mueller,C.G.F., Lane,W.S., Thorner,J. and Hahn,S. (1994) Mot1, a global repressor of RNA polymerase II transcription, inhibits TBP binding to DNA by an ATP-dependent mechanism. Genes Dev., 8, 1920–1934. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
