

IL-17 derived from juxta-articular bone and synovium contributes to joint degradation in rheumatoid arthritis
- PMID: 11299057
- PMCID: PMC30709
- DOI: 10.1186/ar294
IL-17 derived from juxta-articular bone and synovium contributes to joint degradation in rheumatoid arthritis
Abstract
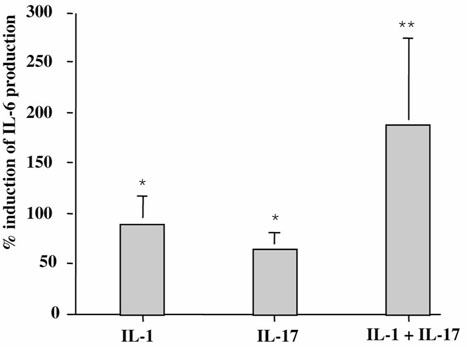
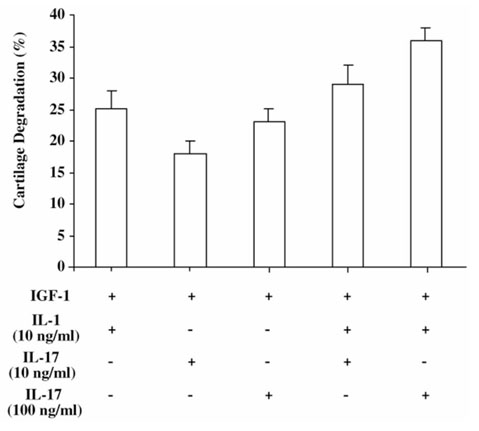
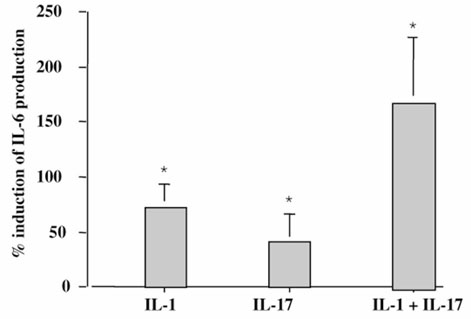
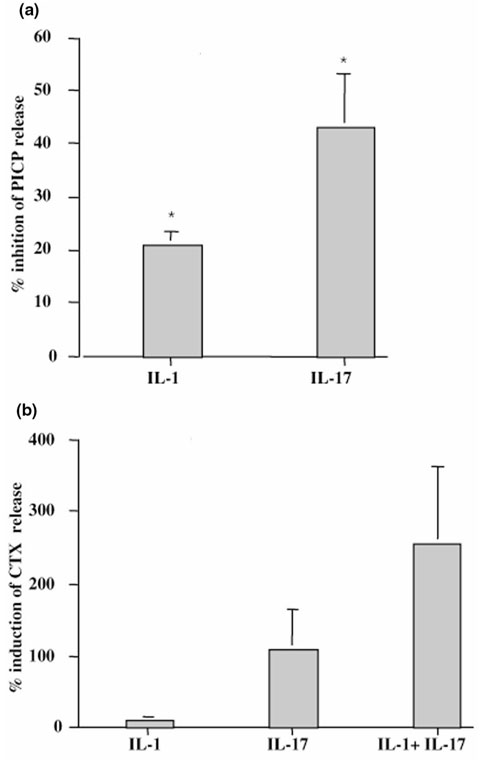
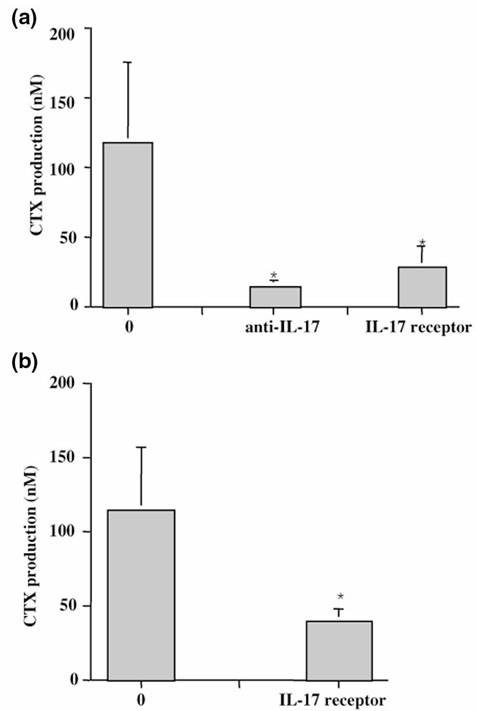
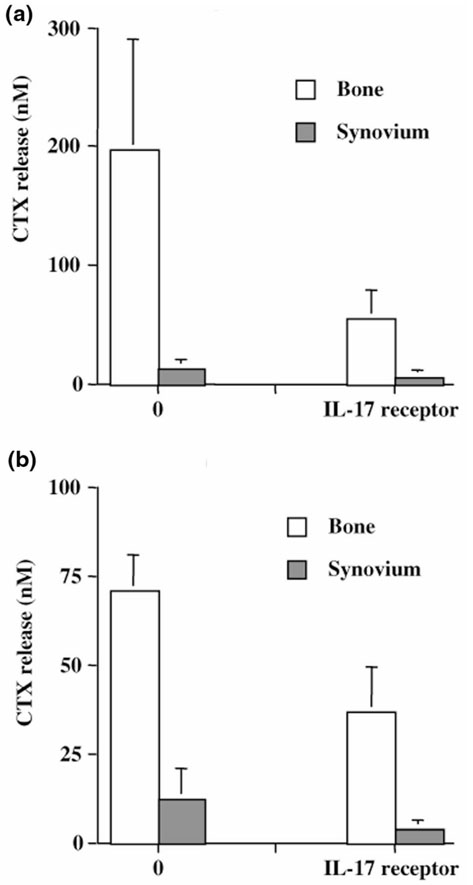
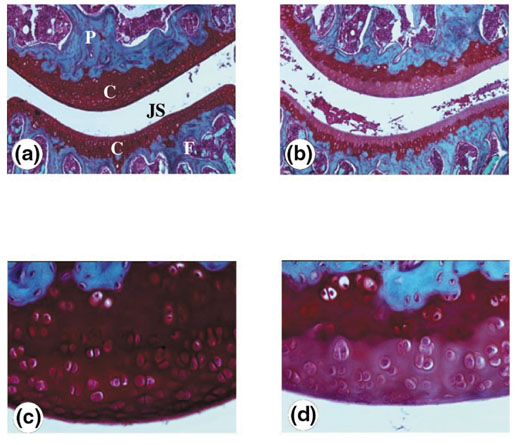
The origin and role of IL-17, a T-cell derived cytokine, in cartilage and bone destruction during rheumatoid arthritis (RA) remain to be clarified. In human ex vivo models, addition of IL-17 enhanced IL-6 production and collagen destruction, and inhibited collagen synthesis by RA synovium explants. On mouse cartilage, IL-17 enhanced cartilage proteoglycan loss and inhibited its synthesis. On human RA bone explants, IL-17 also increased bone resorption and decreased formation. Addition of IL-1 in these conditions increased the effect of IL-17. Blocking of bone-derived endogenous IL-17 with specific inhibitors resulted in a protective inhibition of bone destruction. Conversely, intra-articular administration of IL-17 into a normal mouse joint induced cartilage degradation. In conclusion, the contribution of IL-17 derived from synovium and bone marrow T cells to joint destruction suggests the control of IL-17 for the treatment of RA.
Figures

References
-
- Arend WP. The pathophysiology and treatment of rheumatoid arthritis. Arthritis Rheum. 1997;40:595–597. - PubMed
-
- Chizzolini C, Chicheportiche R, Burger D, Dayer JM. Human Th1 cells preferentially induce interleukin (IL)-1beta while Th2 cells induce IL-1 receptor antagonist production upon cell/cell contact with monocytes. Eur J Immunol. 1997;27:171–177. - PubMed
-
- Chomarat P, Rissoan MC, Pin JJ, Banchereau J, Miossec P. Contribution of IL-1, CD 14, CD 13 in the increased IL-6 production during monocyte synoviocyte interactions. J Immunol. 1995;155:3645–3652. - PubMed
-
- Manolagas SC. Role of cytokines in bone resorption. Bone. 1995;17:63S–67S. - PubMed
-
- Lader CS, Flanagan AM. Prostaglandin E2, interleukin-1a, and tumor necrosis factor-α, increase human osteoblast formation and bone resorption in vitro. Endocrinology. 1998;139:3157–3164. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
