

Molecular characterization of tomato 3-dehydroquinate dehydratase-shikimate:NADP oxidoreductase
- PMID: 11299368
- PMCID: PMC88844
- DOI: 10.1104/pp.125.4.1891
Molecular characterization of tomato 3-dehydroquinate dehydratase-shikimate:NADP oxidoreductase
Abstract
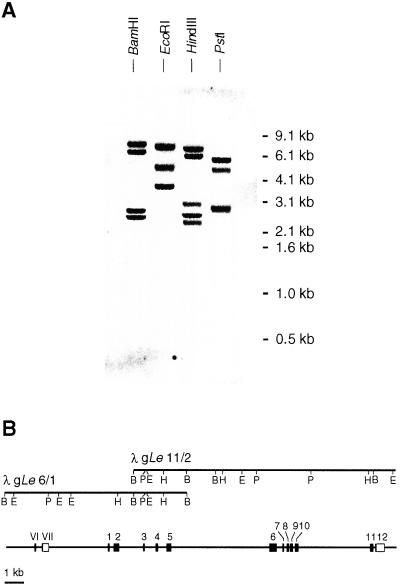
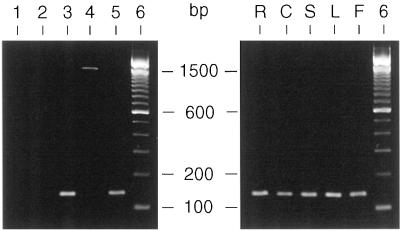
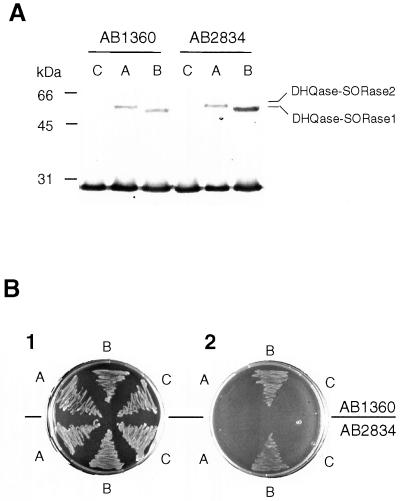
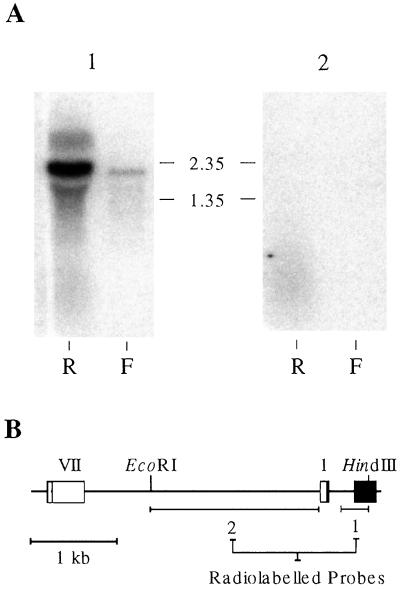
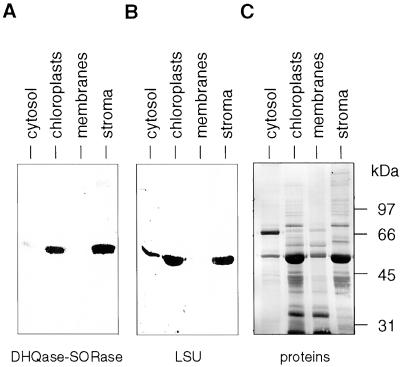
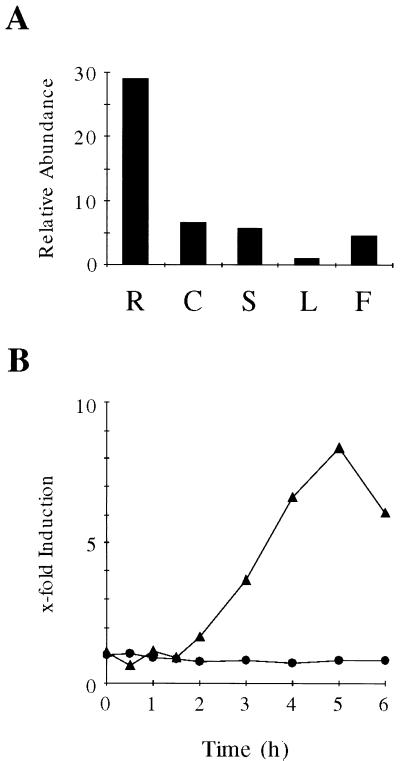
Analysis of cDNAs encoding the bifunctional 3-dehydroquinate dehydratase-shikimate:NADP oxidoreductase (DHQase-SORase) from tomato (Lycopersicon esculentum) revealed two classes of cDNAs that differed by 57 bp within the coding regions, but were otherwise identical. Comparison of these cDNA sequences with the sequence of the corresponding single gene unequivocally proved that the primary transcript is differentially spliced, potentially giving rise to two polypeptides that differ by 19 amino acids. Quantitative real-time polymerase chain reaction revealed that the longer transcript constitutes at most 1% to 2% of DHQase-SORase transcripts. Expression of the respective polypeptides in Escherichia coli mutants lacking the DHQase or the SORase activity gave functional complementation only in case of the shorter polypeptide, indicating that skipping of a potential exon is a prerequisite for the production of an enzymatically active protein. The deduced amino acid sequence revealed that the DHQase-SORase is most likely synthesized as a precursor with a very short (13-amino acid) plastid-specific transit peptide. Like other genes encoding enzymes of the prechorismate pathway in tomato, this gene is elicitor-inducible. Tissue-specific expression resembles the patterns obtained for 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase 2 and dehydroquinate synthase genes. This work completes our studies of the prechorismate pathway in that cDNAs for all seven enzymes (including isozymes) of the prechorismate pathway from tomato have now been characterized.
Figures

References
-
- Aebi M, Hornig H, Padgett RA, Reiser J, Weissmann C. Sequence requirements for splicing of higher eukaryotic nuclear pre-mRNA. Cell. 1986;47:555–565. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. Chichester, UK: John Wiley & Sons; 1994.
-
- Bentley R. The shikimate pathway: a metabolic tree with many branches. Crit Rev Biochem Mol Biol. 1990;25:307–384. - PubMed
-
- Bischoff M, Rösler J, Raesecke H-R, Görlach J, Amrhein N, Schmid J. Cloning of a cDNA encoding a 3-dehydroquinate synthase from a higher plant, and analysis of the organ-specific and elicitor-induced expression of the corresponding gene. Plant Mol Biol. 1996;31:69–76. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
