

Identification of a chlamydial protease-like activity factor responsible for the degradation of host transcription factors
- PMID: 11304554
- PMCID: PMC2193410
- DOI: 10.1084/jem.193.8.935
Identification of a chlamydial protease-like activity factor responsible for the degradation of host transcription factors
Abstract
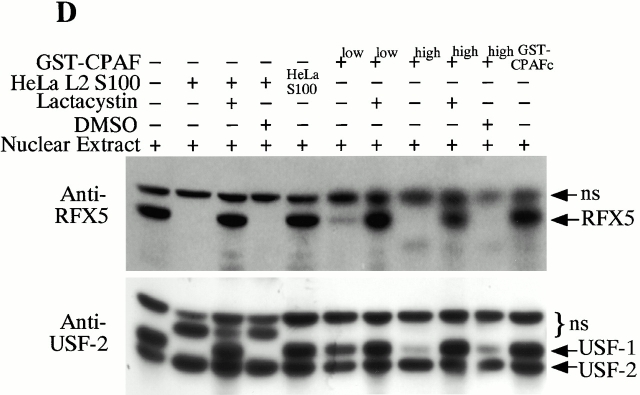
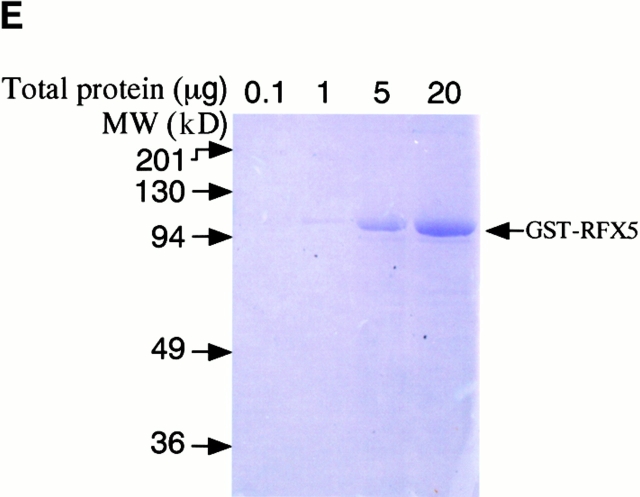
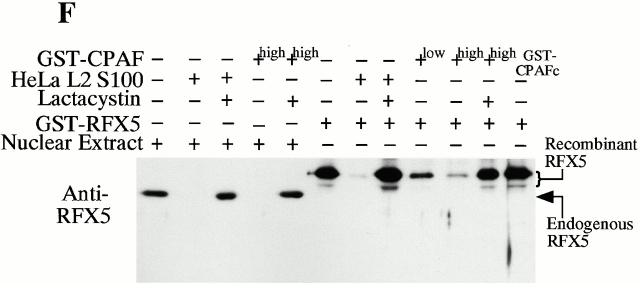
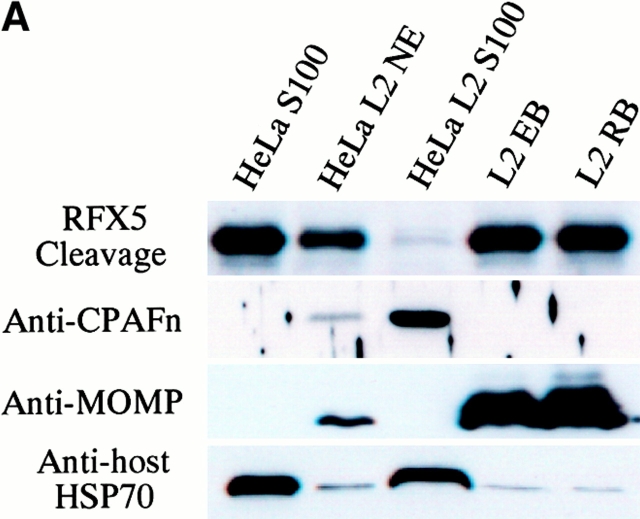
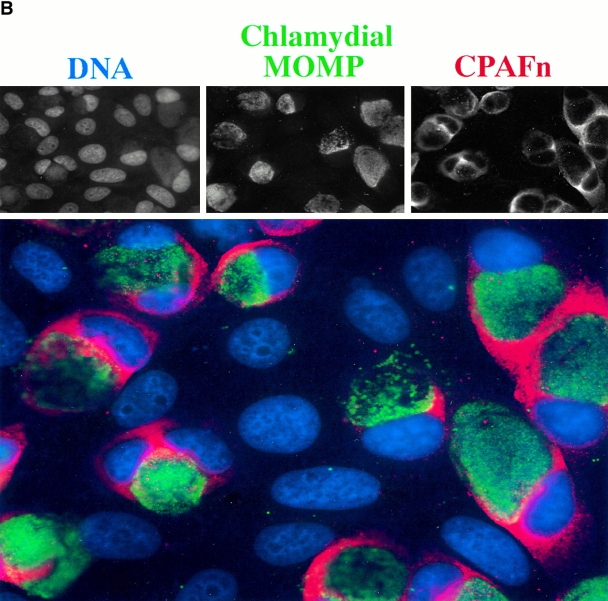
Microbial pathogens have been selected for the capacity to evade or manipulate host responses in order to survive after infection. Chlamydia, an obligate intracellular pathogen and the causative agent for many human diseases, can escape T lymphocyte immune recognition by degrading host transcription factors required for major histocompatibility complex (MHC) antigen expression. We have now identified a chlamydial protease- or proteasome-like activity factor (CPAF) that is secreted into the host cell cytosol and that is both necessary and sufficient for the degradation of host transcription factors RFX5 and upstream stimulation factor 1 (USF-1). The CPAF gene is highly conserved among chlamydial strains, but has no significant overall homology with other known genes. Thus, CPAF represents a unique secreted protein produced by an obligate intracellular bacterial pathogen to interfere with effective host adaptive immunity.
Figures
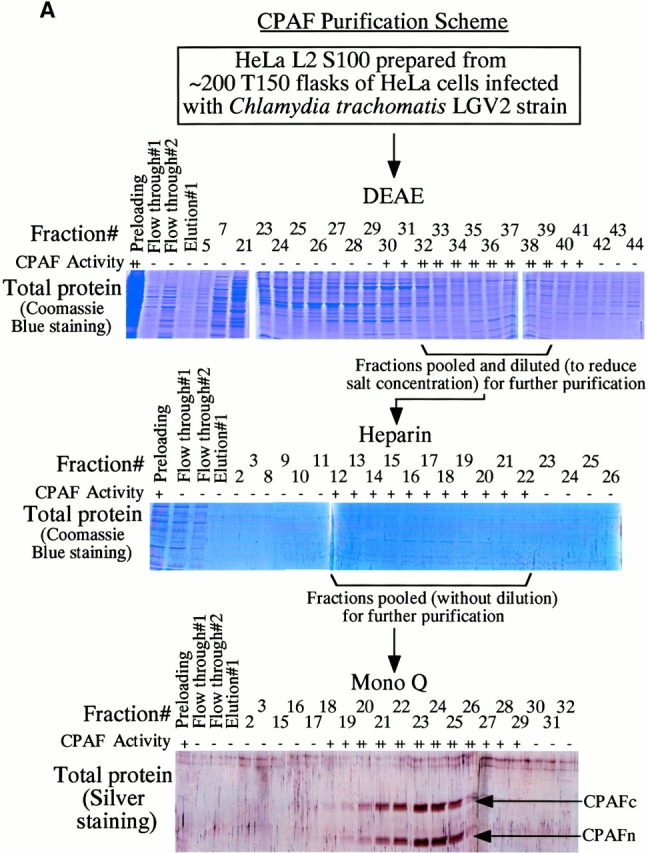
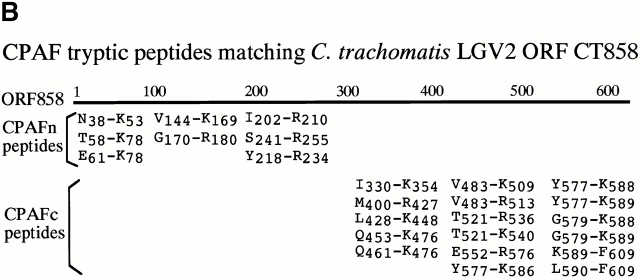
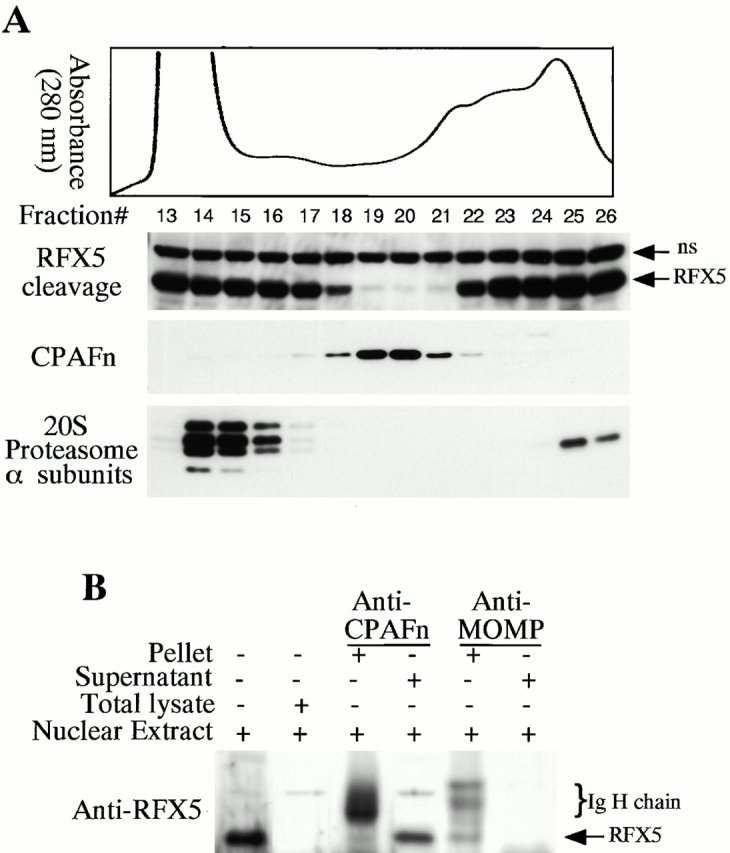
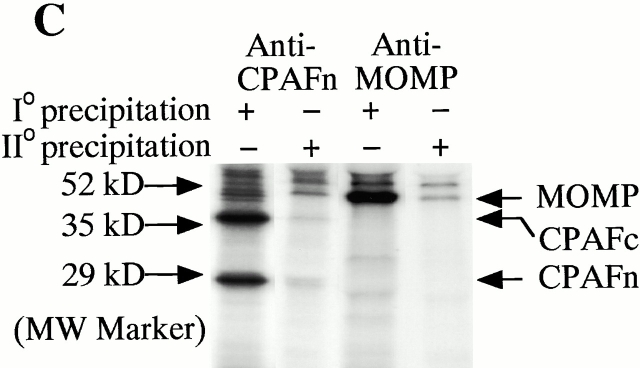
References
-
- Rasmussen S.J., Eckmann L., Quayle A.J., Shen L., Zhang Y.X., Anderson D.J., Fierer J., Stephens R.S., Kagnoff M.F. Secretion of proinflammatory cytokines by epithelial cells in response to Chlamydia infection suggests a central role for epithelial cells in chlamydial pathogenesis. J. Clin. Invest. 1997;99:77–87. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
