

Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)
- PMID: 11306629
- PMCID: PMC6762533
- DOI: 10.1523/JNEUROSCI.21-08-02768.2001
Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)
Abstract
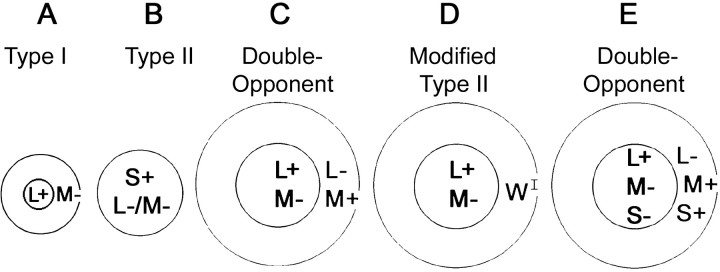
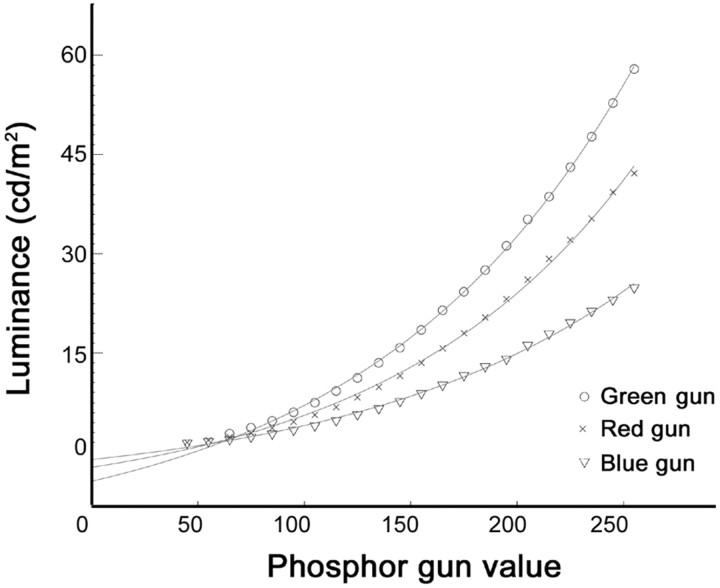
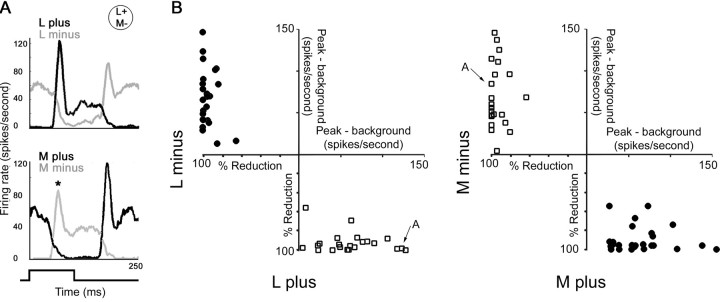
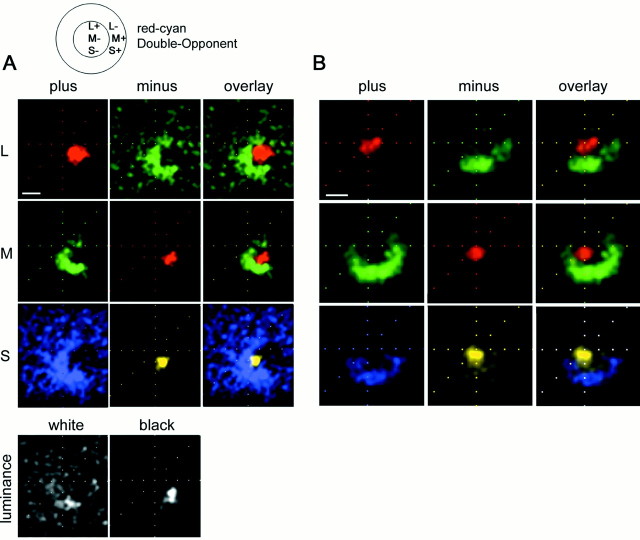
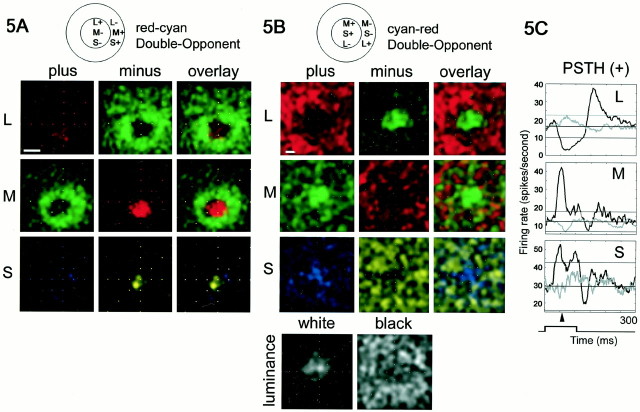
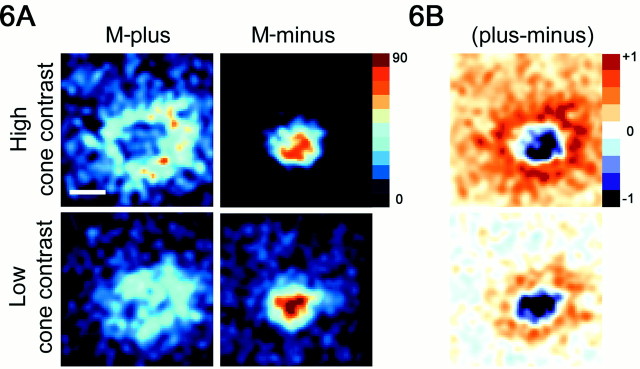
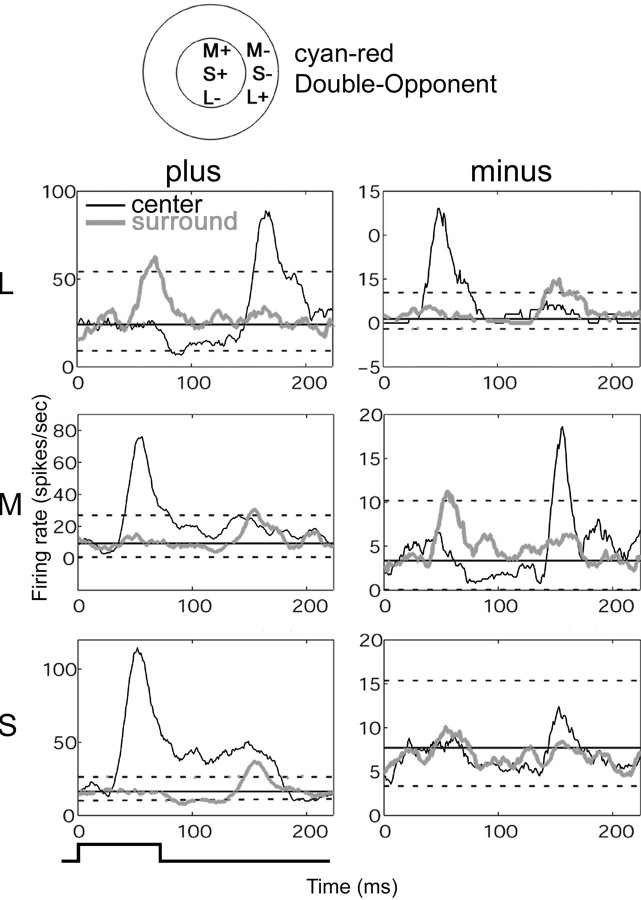
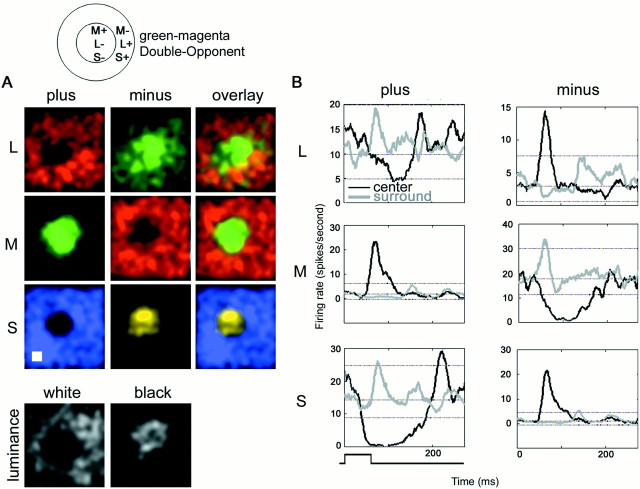
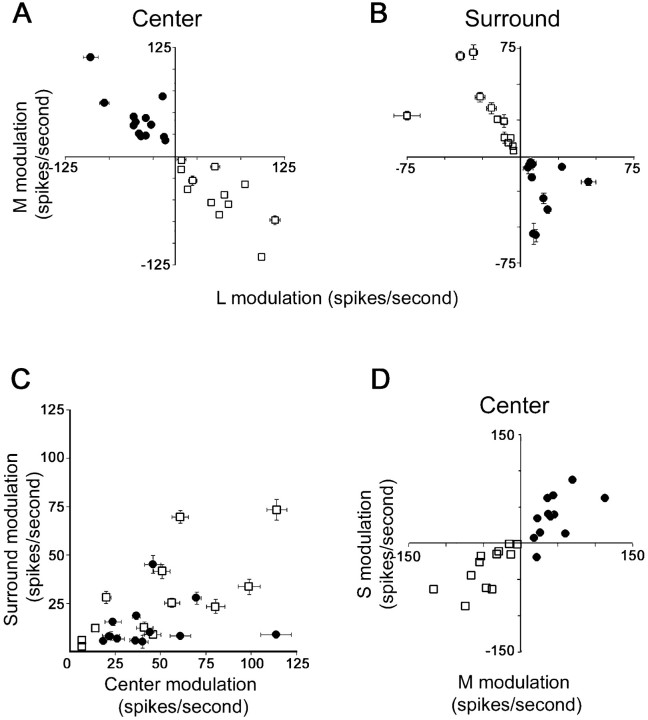
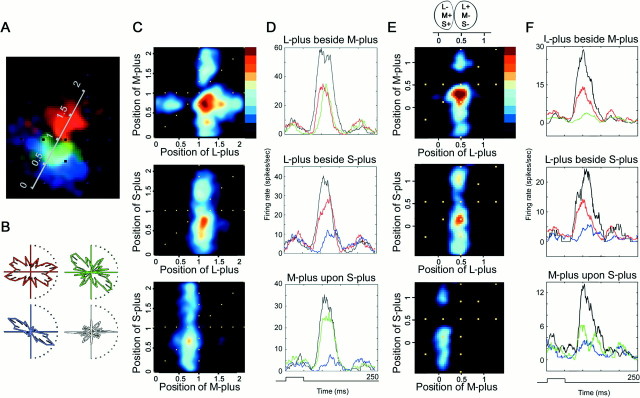
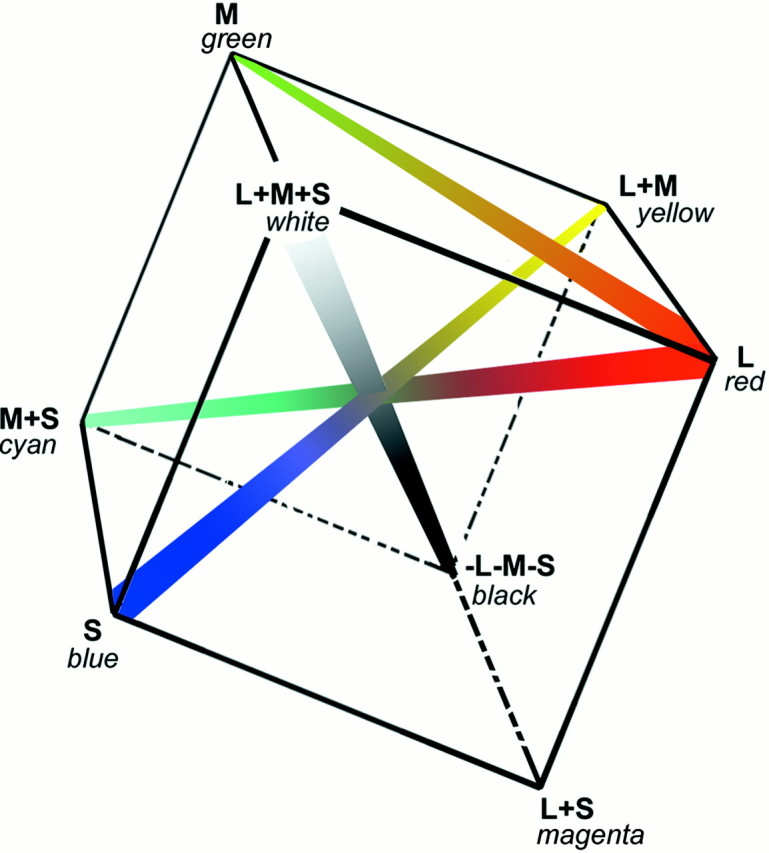
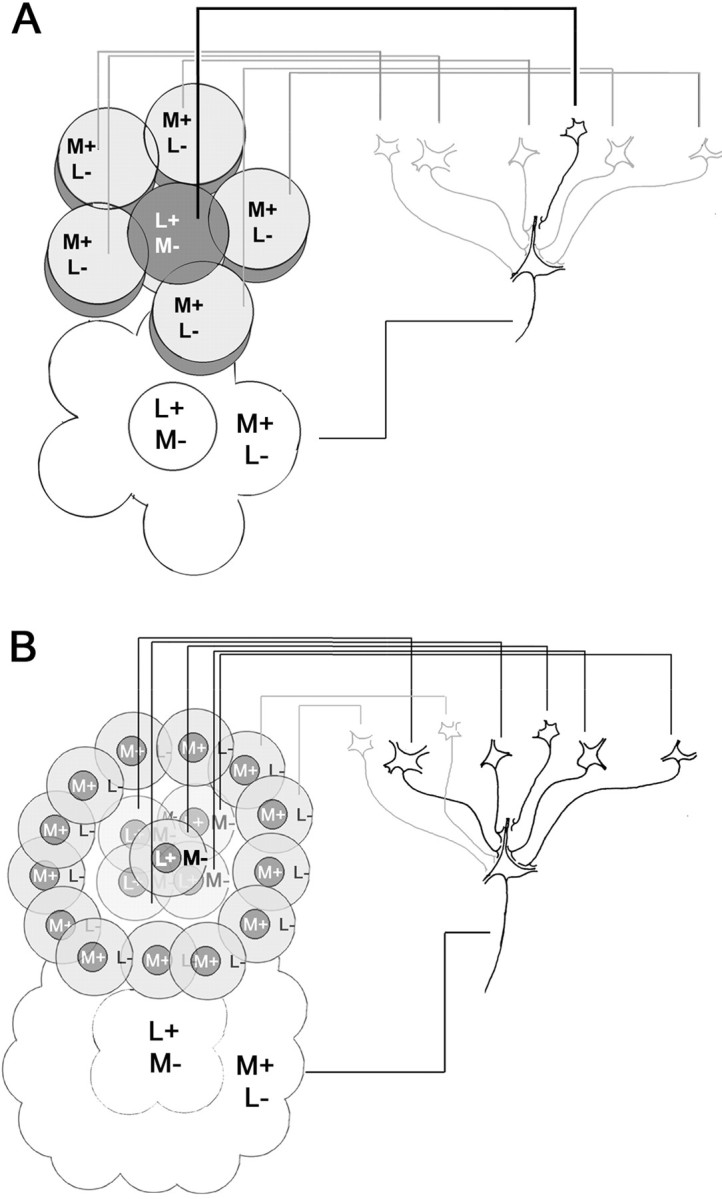
The spatial structure of color cell receptive fields is controversial. Here, spots of light that selectively modulate one class of cones (L, M, or S, or loosely red, green, or blue) were flashed in and around the receptive fields of V-1 color cells to map the spatial structure of the cone inputs. The maps generated using these cone-isolating stimuli and an eye-position-corrected reverse correlation technique produced four findings. First, the receptive fields were Double-Opponent, an organization of spatial and chromatic opponency critical for color constancy and color contrast. Optimally stimulating both center and surround subregions with adjacent red and green spots excited the cells more than stimulating a single subregion. Second, red-green cells responded in a luminance-invariant way. For example, red-on-center cells were excited equally by a stimulus that increased L-cone activity (appearing bright red) and by a stimulus that decreased M-cone activity (appearing dark red). This implies that the opponency between L and M is balanced and argues that these cells are encoding a single chromatic axis. Third, most color cells responded to stimuli of all orientations and had circularly symmetric receptive fields. Some cells, however, showed a coarse orientation preference. This was reflected in the receptive fields as oriented Double-Opponent subregions. Fourth, red-green cells often responded to S-cone stimuli. Responses to M- and S-cone stimuli usually aligned, suggesting that these cells might be red-cyan. In summary, red-green (or red-cyan) cells, along with blue-yellow and black-white cells, establish three chromatic axes that are sufficient to describe all of color space.
Figures
References
-
- Albers J. Interaction of color, pp 20–21. Yale UP; New Haven, CT: 1963.
-
- Anderson JC, Martin KA, Whitteridge D. Form, function, and intracortical projections of neurons in the striate cortex of the monkey Macacus nemestrinus. Cereb Cortex. 1993;3:412–420. - PubMed
-
- Calkins DJ, Sterling P. Evidence that circuits for spatial and color vision segregate at the first retinal synapse. Neuron. 1999;24:313–321. - PubMed
-
- Cavonius CR, Schumacher AW. Human visual acuity measured with colored test objects. Science. 1966;152:1276–1277. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources