

The preferred stoichiometry of c subunits in the rotary motor sector of Escherichia coli ATP synthase is 10
- PMID: 11320246
- PMCID: PMC33147
- DOI: 10.1073/pnas.081424898
The preferred stoichiometry of c subunits in the rotary motor sector of Escherichia coli ATP synthase is 10
Abstract
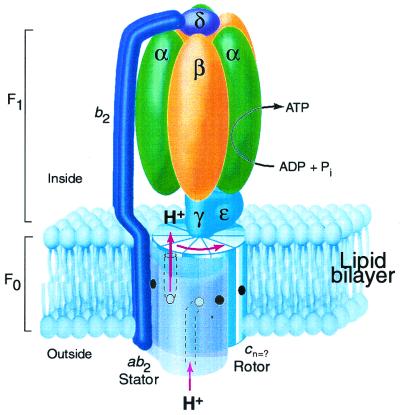
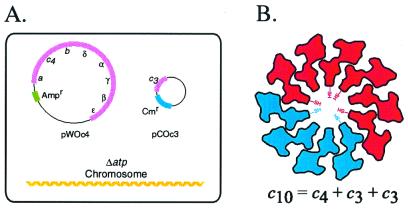
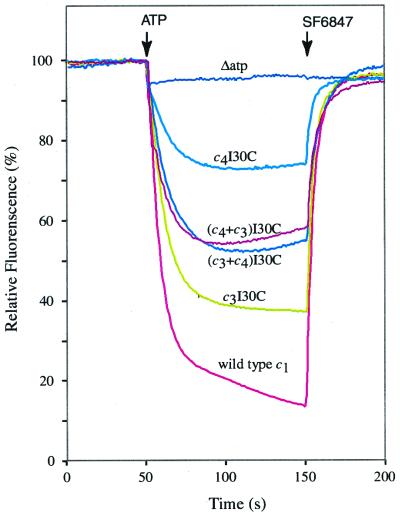
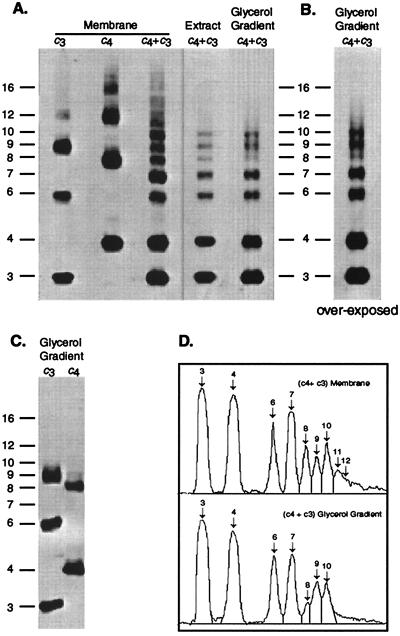
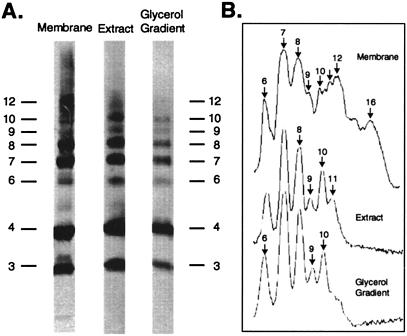
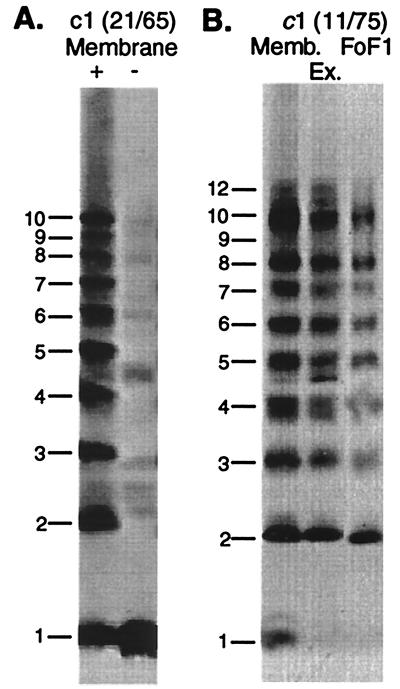
The stoichiometry of c subunits in the H(+)-transporting F(o) rotary motor of ATP synthase is uncertain, the most recent suggestions varying from 10 to 14. The stoichiometry will determine the number of H(+) transported per ATP synthesized and will directly relate to the P/O ratio of oxidative phosphorylation. The experiments described here show that the number of c subunits in functional complexes of F(o)F(1) ATP synthase from Escherichia coli can be manipulated, but that the preferred number is 10. Mixtures of genetically fused cysteine-substituted trimers (c(3)) and tetramers (c(4)) of subunit c were coexpressed and the c subunits crosslinked in the plasma membrane. Prominent products corresponding to oligomers of c(7) and c(10) were observed in the membrane and purified F(o)F(1) complex, indicating that the c(10) oligomer formed naturally. Oligomers larger than c(10) were also observed in the membrane fraction of cells expressing c(3) or c(4) individually, or in cells coexpressing c(3) and c(4) together, but these larger oligomers did not copurify with the functional F(o)F(1) complex and were concluded to be aberrant products of assembly in the membrane.
Figures
References
-
- Boyer P D. Annu Rev Biochem. 1997;66:717–749. - PubMed
-
- Noji H, Yasuda R, Yoshida M, Kinosita K., Jr Nature (London) 1997;386:299–302. - PubMed
-
- Sabert D, Engelbrecht S, Junge W. Nature (London) 1996;381:623–625. - PubMed
-
- Sambongi Y, Iko Y, Tanabe M, Omote H, Iwasmoto-Kihara A, Ueda I, Yanagida T, Wada Y, Futai M. Science. 1999;286:1722–1724. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
