

Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes
- PMID: 11320249
- PMCID: PMC33171
- DOI: 10.1073/pnas.091577598
Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes
Abstract
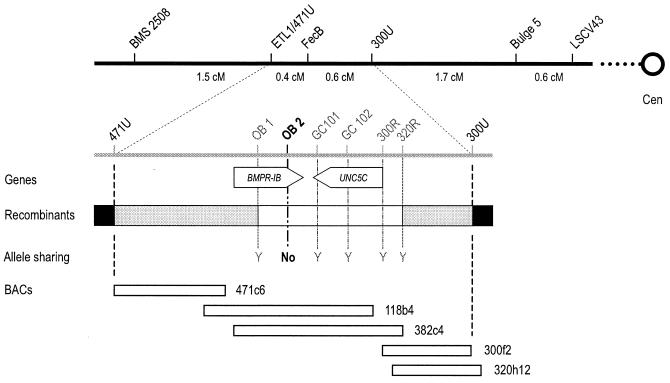
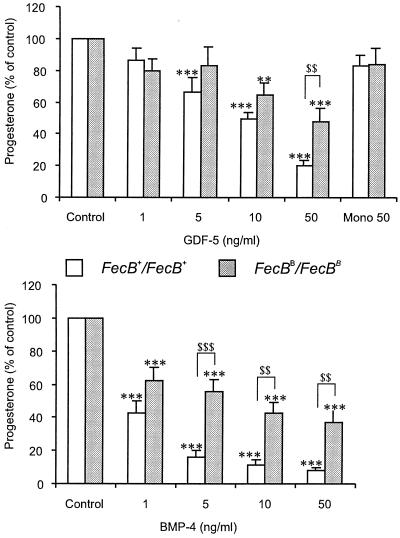
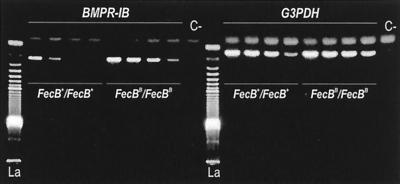
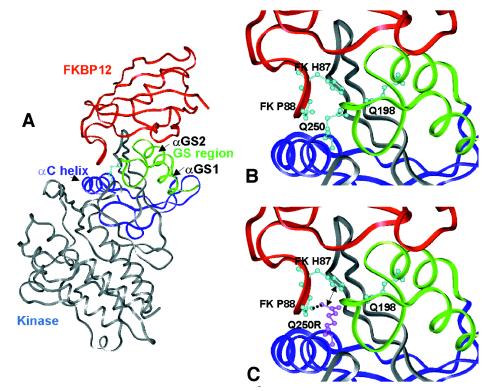
Ewes from the Booroola strain of Australian Mérino sheep are characterized by high ovulation rate and litter size. This phenotype is due to the action of the FecB(B) allele of a major gene named FecB, as determined by statistical analysis of phenotypic data. By genetic analysis of 31 informative half-sib families from heterozygous sires, we showed that the FecB locus is situated in the region of ovine chromosome 6 corresponding to the human chromosome 4q22-23 that contains the bone morphogenetic protein receptor IB (BMPR-IB) gene encoding a member of the transforming growth factor-beta (TGF-beta) receptor family. A nonconservative substitution (Q249R) in the BMPR-IB coding sequence was found to be associated fully with the hyperprolificacy phenotype of Booroola ewes. In vitro, ovarian granulosa cells from FecB(B)/FecB(B) ewes were less responsive than granulosa cells from FecB(+)/FecB(+) ewes to the inhibitory effect on steroidogenesis of GDF-5 and BMP-4, natural ligands of BMPR-IB. It is suggested that in FecB(B)/FecB(B) ewes, BMPR-IB would be inactivated partially, leading to an advanced differentiation of granulosa cells and an advanced maturation of ovulatory follicles.
Figures

References
-
- Davis G H, Montgomery G W, Allision A J, Kelly R W, Bray A R. N Z J Agric Res. 1982;25:525–529.
-
- Piper L R, Bindon B M, Davis G H. In: Genetics of Reproduction in Sheep. Land R B, Robinson D W, editors. London: Butterworths; 1985. pp. 115–125.
-
- McNatty K P, Henderson K M. J Steroid Biochem. 1987;27:365–373. - PubMed
-
- Montgomery G W. Rev Reprod. 2000;5:25–37. - PubMed
-
- Montgomery G W, Crawford A M, Penty J M, Dodds K G, Ede A J, Henry H M, Pierson C A, Lord E A, Galloway S M, Schmack A E, et al. Nat Genet. 1993;4:410–414. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
