

Modular construction for function of a ribonucleoprotein enzyme: the catalytic domain of Bacillus subtilis RNase P complexed with B. subtilis RNase P protein
- PMID: 11328872
- PMCID: PMC37254
- DOI: 10.1093/nar/29.9.1892
Modular construction for function of a ribonucleoprotein enzyme: the catalytic domain of Bacillus subtilis RNase P complexed with B. subtilis RNase P protein
Abstract
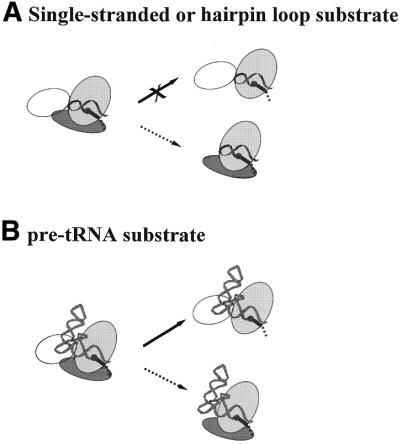
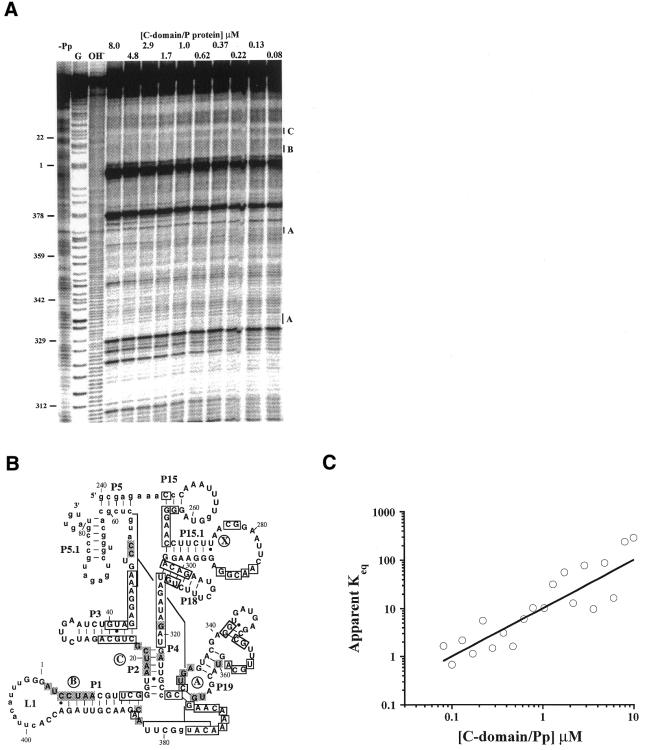
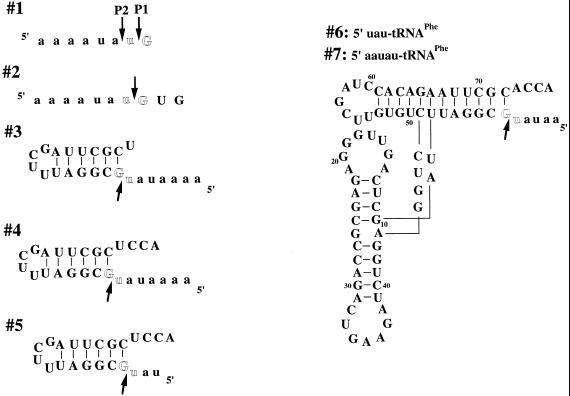
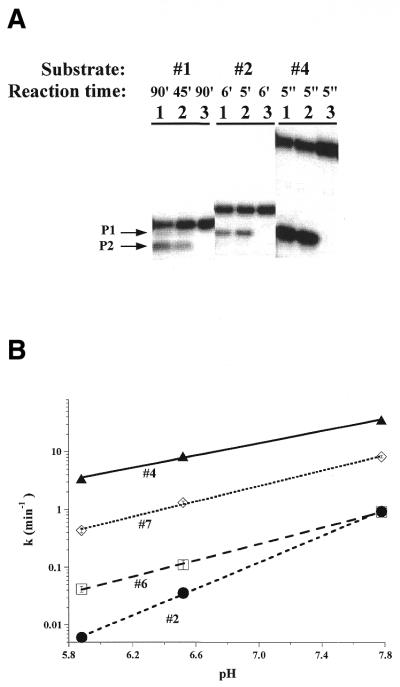
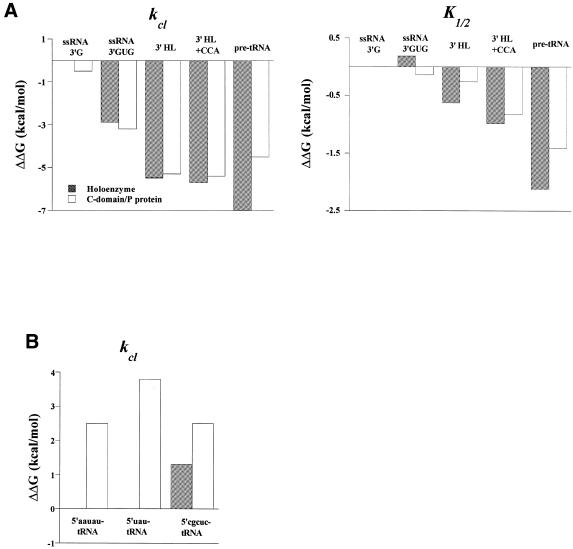
The bacterial RNase P holoenzyme catalyzes the formation of the mature 5'-end of tRNAs and is composed of an RNA and a protein subunit. Among the two folding domains of the RNase P RNA, the catalytic domain (C-domain) contains the active site of this ribozyme. We investigated specific binding of the Bacillus subtilis C-domain with the B.subtilis RNase P protein and examined the catalytic activity of this C-domain-P protein complex. The C-domain forms a specific complex with the P protein with a binding constant of approximately 0.1 microM. The C-domain-P protein complex and the holoenzyme are equally efficient in cleaving single-stranded RNA (approximately 0.9 min(-1) at pH 7.8) and substrates with a hairpin-loop 3' to the cleavage site (approximately 40 min(-1)). The holoenzyme reaction is much more efficient with a pre-tRNA substrate, binding at least 100-fold better and cleaving 10-500 times more efficiently. These results demonstrate that the RNase P holoenzyme is functionally constructed in three parts. The catalytic domain alone contains the active site, but has little specificity and affinity for most substrates. The specificity and affinity for the substrate is generated by either the specificity domain of RNase P RNA binding to a T stem-loop-like hairpin or RNase P protein binding to a single-stranded RNA. This modular construction may be exploited to obtain RNase P-based ribonucleoprotein complexes with altered substrate specificity.
Figures
References
-
- Frank D.N. and Pace,N.R. (1998) Ribonuclease P: unity and diversity in a tRNA processing ribozyme. Annu. Rev. Biochem., 67, 153–180. - PubMed
-
- Altman S. and Kirsebom,L. (1999) Ribonuclease P. In Gesteland,R.F., Cech,T.R. and Atkins,J.F. (eds), The RNA World. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 351–380.
-
- Massire C., Jaeger,L. and Westhof,E. (1998) Derivation of the three-dimensional architecture of bacterial ribonuclease P RNAs from comparative sequence analysis. J. Mol. Biol., 279, 773–793. - PubMed
-
- Fang X., Pan,T. and Sosnick,T.R. (1999) A thermodynamic framework and cooperativity in the tertiary folding of a Mg2+-dependent ribozyme. Biochemistry, 38, 16840–16846. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
