

Rho-kinase--mediated contraction of isolated stress fibers
- PMID: 11331307
- PMCID: PMC2190572
- DOI: 10.1083/jcb.153.3.569
Rho-kinase--mediated contraction of isolated stress fibers
Abstract
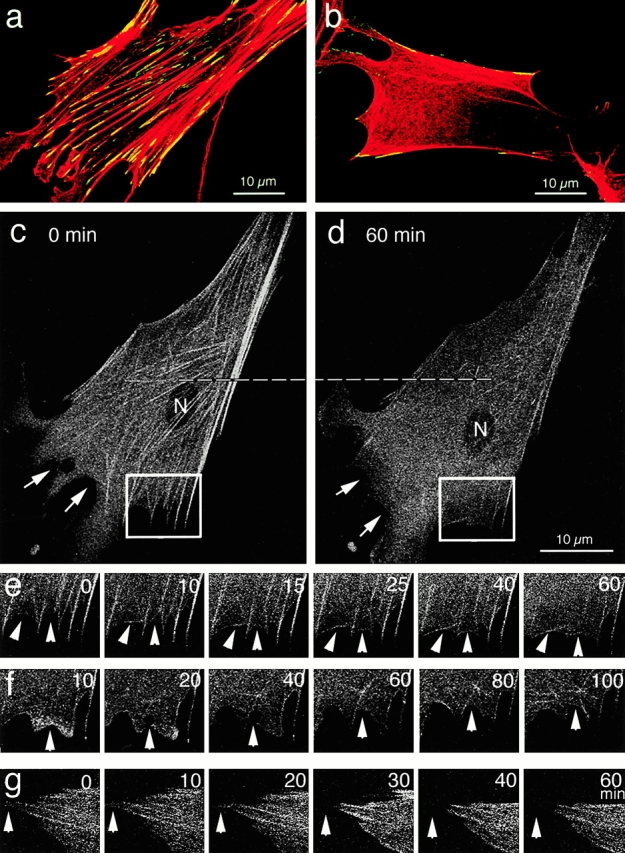
It is widely accepted that actin filaments and the conventional double-headed myosin interact to generate force for many types of nonmuscle cell motility, and that this interaction occurs when the myosin regulatory light chain (MLC) is phosphorylated by MLC kinase (MLCK) together with calmodulin and Ca(2+). However, recent studies indicate that Rho-kinase is also involved in regulating the smooth muscle and nonmuscle cell contractility. We have recently isolated reactivatable stress fibers from cultured cells and established them as a model system for actomyosin-based contraction in nonmuscle cells. Here, using isolated stress fibers, we show that Rho-kinase mediates MLC phosphorylation and their contraction in the absence of Ca(2+). More rapid and extensive stress fiber contraction was induced by MLCK than was by Rho-kinase. When the activity of Rho-kinase but not MLCK was inhibited, cells not only lost their stress fibers and focal adhesions but also appeared to lose cytoplasmic tension. Our study suggests that actomyosin-based nonmuscle contractility is regulated by two kinase systems: the Ca(2+)-dependent MLCK and the Rho-kinase systems. We propose that Ca(2+) is used to generate rapid contraction, whereas Rho-kinase plays a major role in maintaining sustained contraction in cells.
Figures

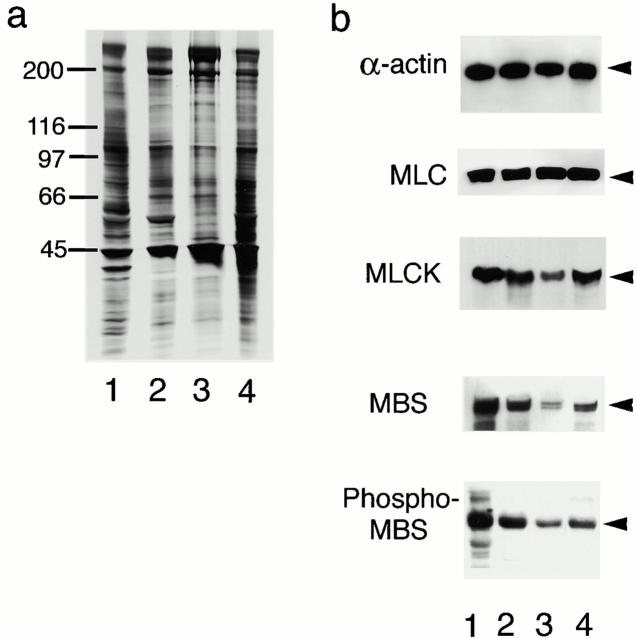
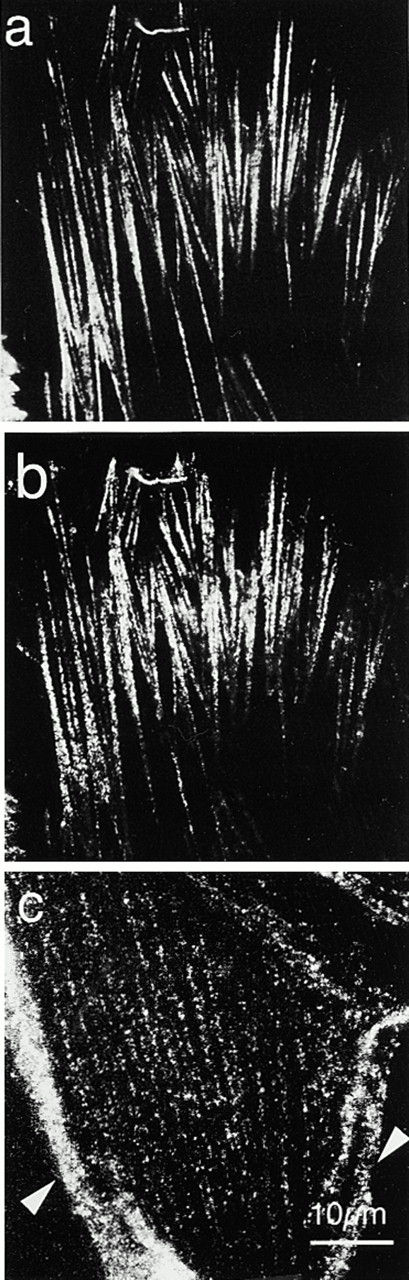
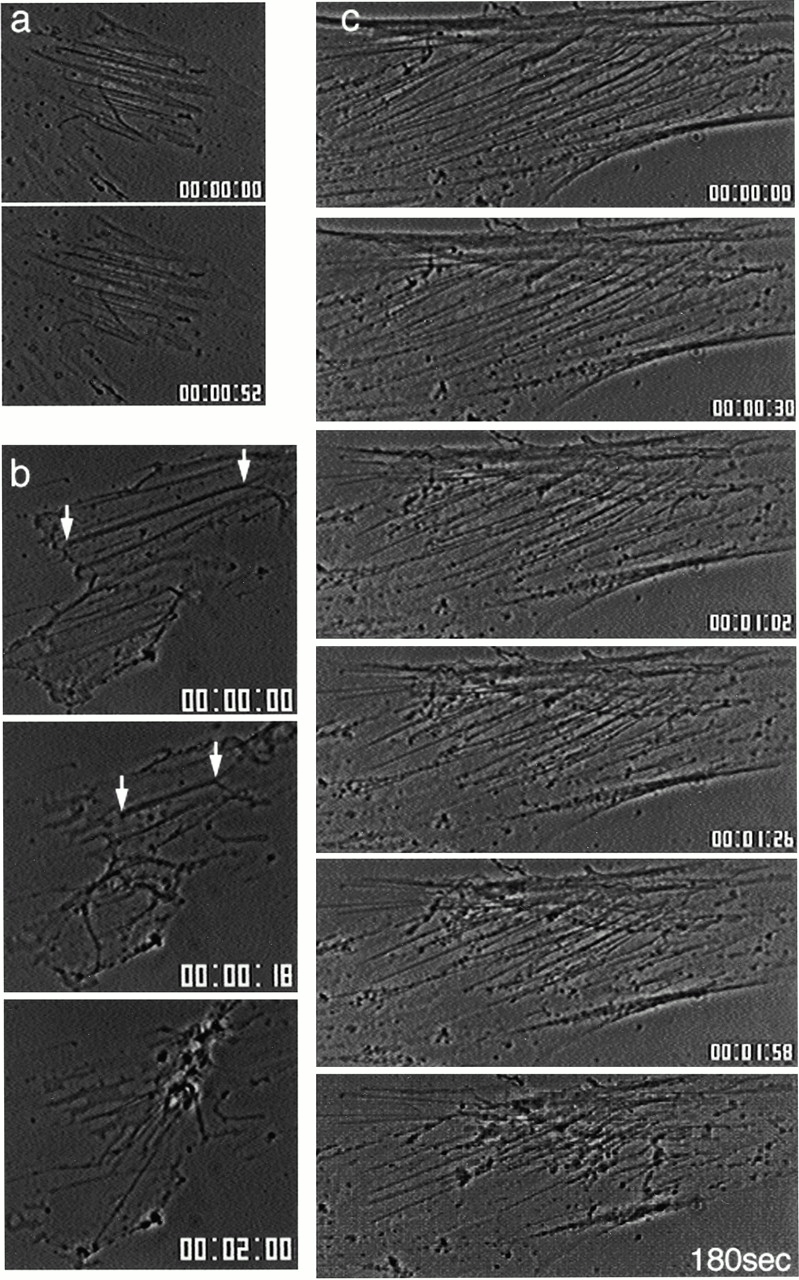
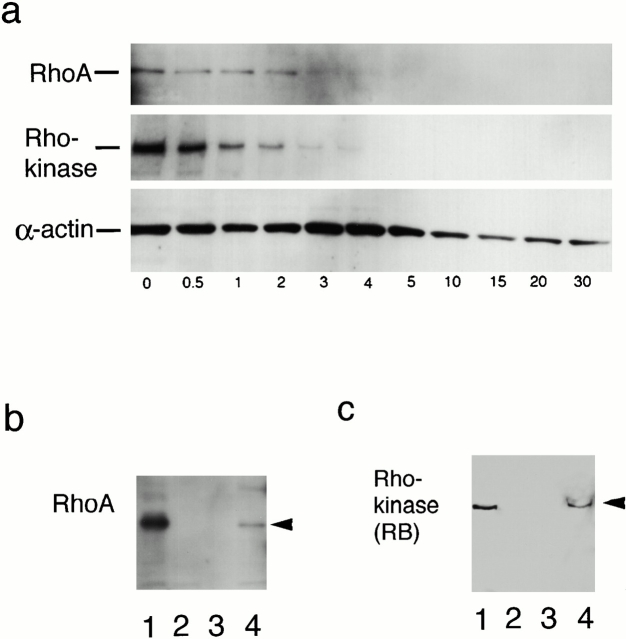
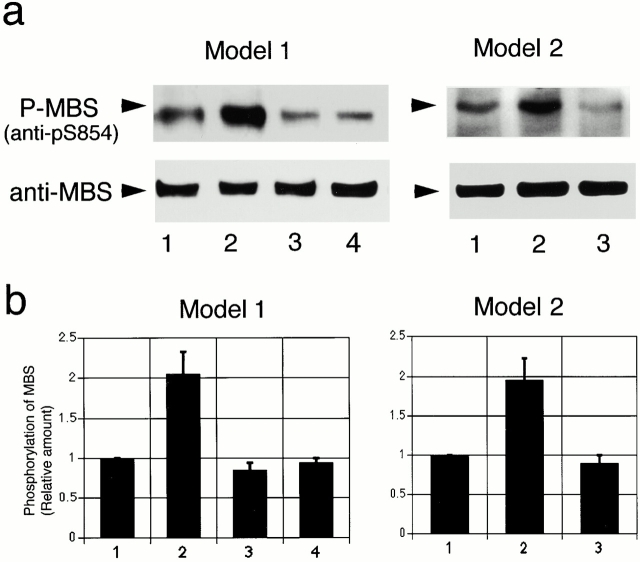
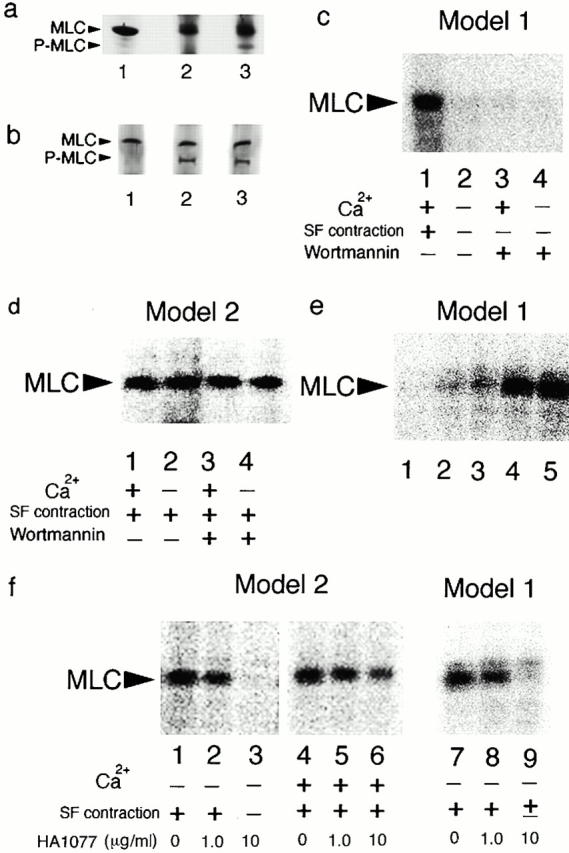
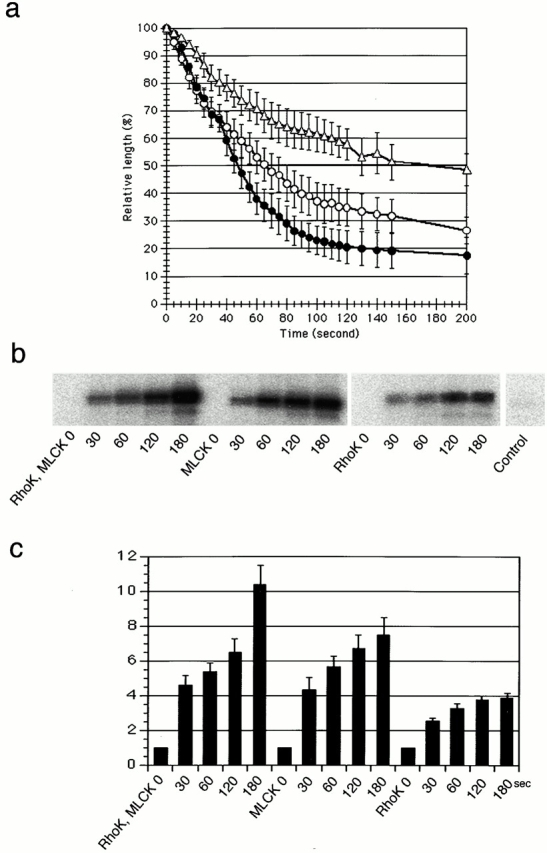
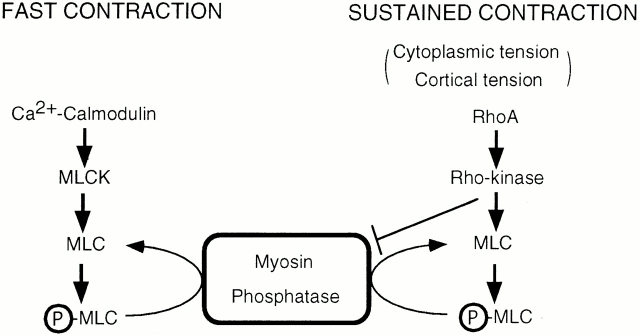
References
-
- Alessi D., MacDougall L.K., Sola M.M., Ikebe M., Cohen P. The control of protein phosphatase-1 by targeting subunits. The major myosin phosphatase in avian smooth muscle is a novel form of protein phosphatase-1. Eur. J. Biochem. 1992;210:1023–1035. - PubMed
-
- Amano M., Ito M., Kimura K., Fukata Y., Chihara K., Nakano T., Matsuura Y., Kaibuchi K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase) J. Biol. Chem. 271 1996. 20246 20249a - PubMed
-
- Amano M., Mukai H., Ono Y., Chihara K., Matsui T., Hamajima Y., Okawa K., Iwamatsu A., Kaibuchi K. Identification of a putative target for Rho as the serine-threonin kinase protein kinase N Science 271 1996. 648 650b - PubMed
-
- Amano M., Chihara K., Kimura K., Fukata Y., Nakamura N., Matsuura Y., Kaibuchi K. Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science. 1997;275:1308–1311. - PubMed
-
- Amano M., Chihara K., Nakamura N., Fukata Y., Yano T., Shibata M., Ikebe M., Kaibuchi K. Myosin II activation promotes neurite retraction during the action of Rho and Rho-kinase. Genes Cells. 1998;3:177–188. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
