

Ezrin-dependent promotion of glioma cell clonogenicity, motility, and invasion mediated by BCL-2 and transforming growth factor-beta2
- PMID: 11331365
- PMCID: PMC6762489
- DOI: 10.1523/JNEUROSCI.21-10-03360.2001
Ezrin-dependent promotion of glioma cell clonogenicity, motility, and invasion mediated by BCL-2 and transforming growth factor-beta2
Abstract
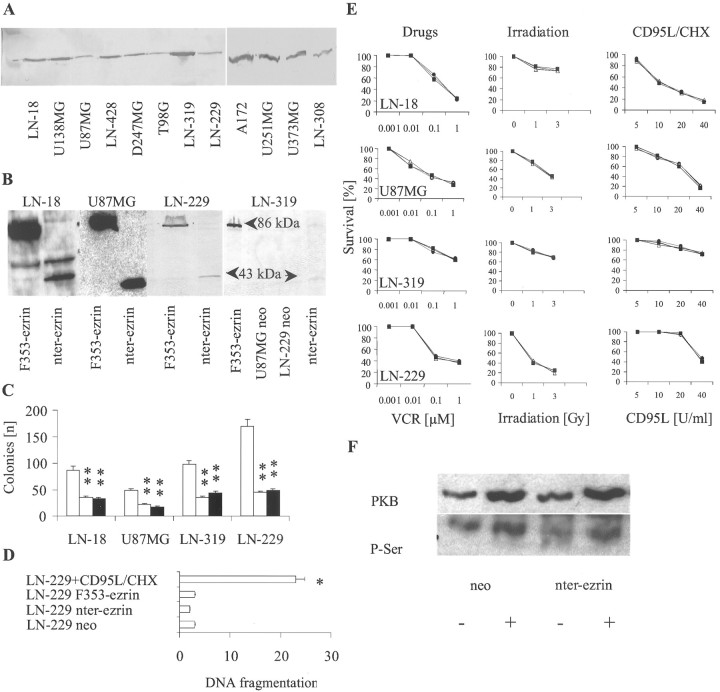
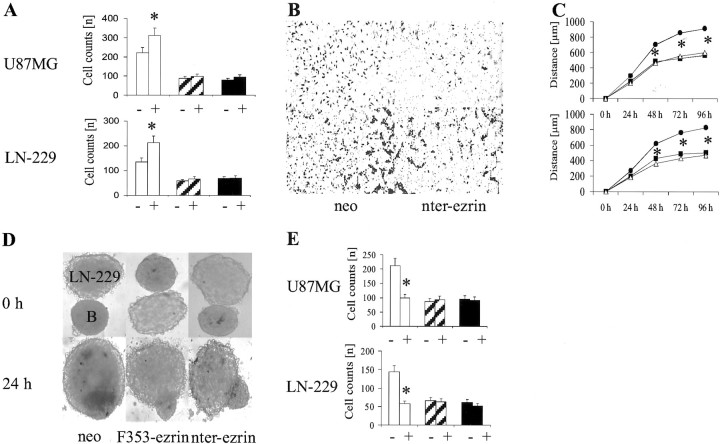
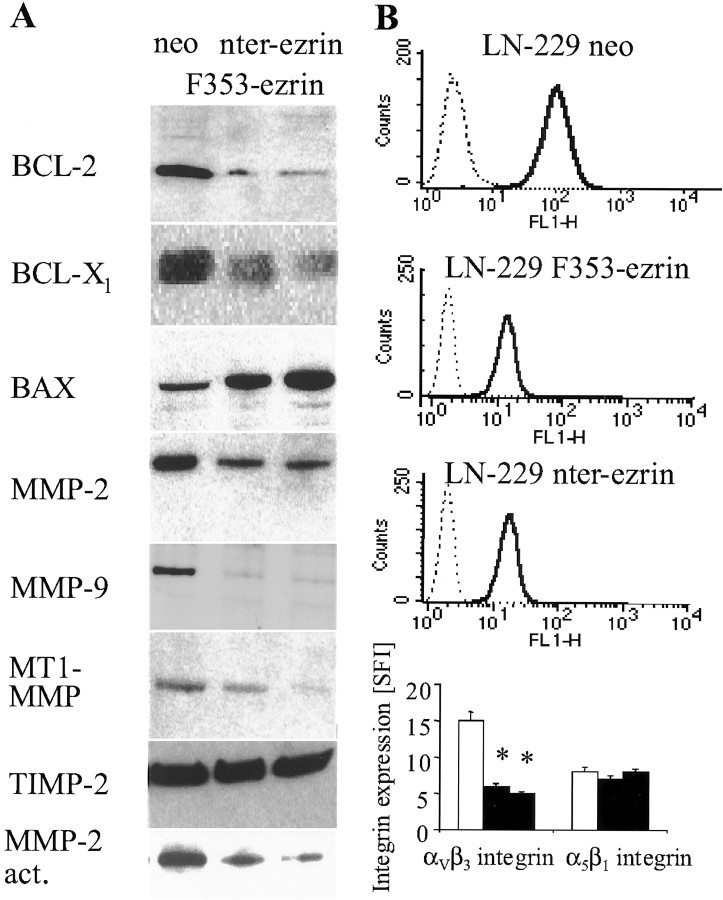
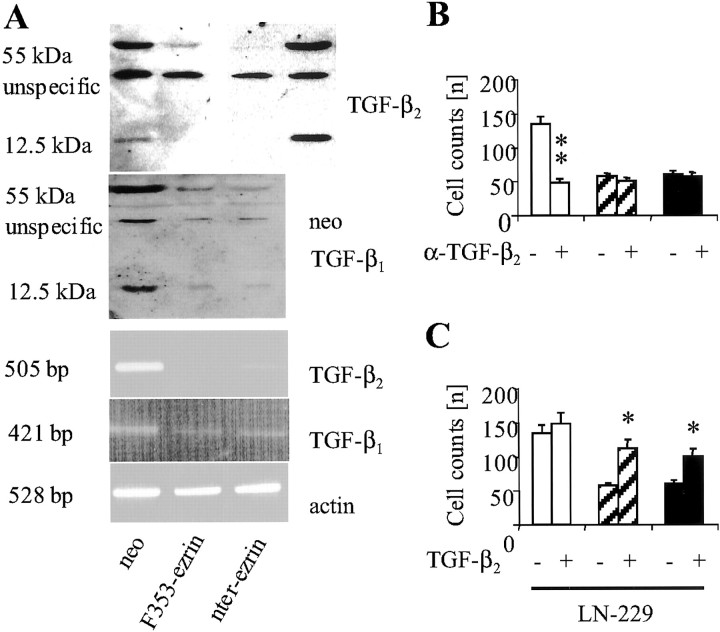
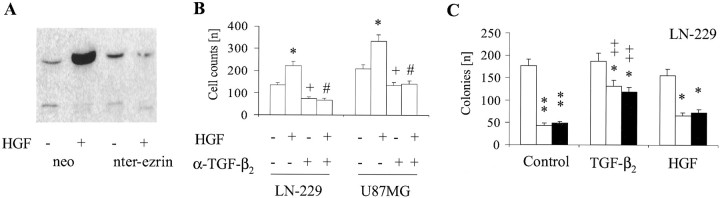
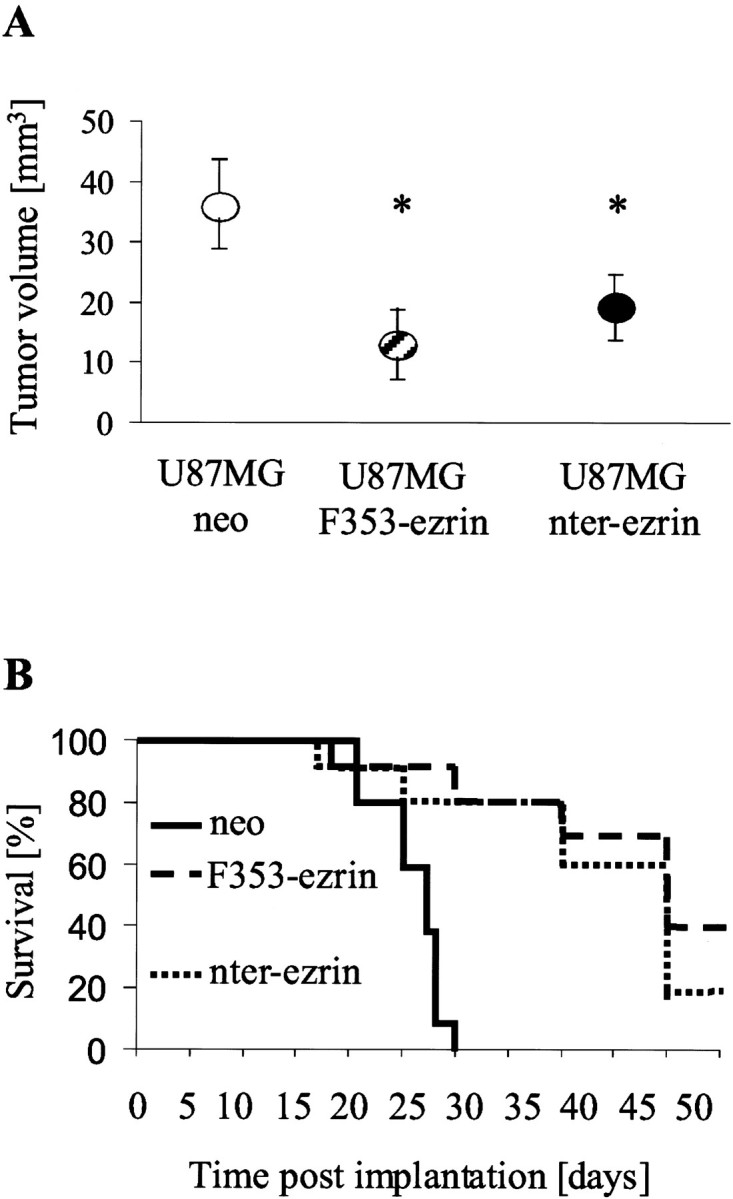
Ezrin belongs to the ezrin-radixin-moesin family proteins, which cross-link actin cytoskeleton and plasma membrane. Malignant glioma cells are paradigmatic for their strong migratory and invasive properties. Here, we report that the expression of dominant-negative ezrins inhibits clonogenicity, migration, and invasiveness of human malignant glioma cells. Furthermore, dominant-negative ezrins block hepatocyte growth factor (HGF)-mediated stimulation of clonogenicity and migration, without altering HGF-induced protein kinase B/Akt and focal adhesion kinase phosphorylation. Glioma cells expressing dominant-negative ezrins exhibit a shift of the BCL-2/BAX rheostat toward apoptosis, reduced alpha(V)beta(3) integrin expression and reduced matrix metalloproteinase (MMP) expression and activity. These changes are associated with a dramatic loss of transforming growth factor beta(2) (TGF-beta(2)) release. Exogenous supplementation of TGF-beta(2) overcomes the inhibitory effects of dominant-negative ezrins on migration and clonogenicity. A neutralizing TGF-beta(2) antibody mimics the effects of dominant-negative ezrins on clonogenicity and migration. Exogenous HGF markedly induces TGF-beta(2) protein levels, and a neutralizing TGF-beta(2) antibody abolishes the HGF-mediated increase in glioma cell motility. Finally, TGF-beta(2) does not modulate BCL-2 or BAX expression, but BCL-2 gene transfer increases the levels of latent and active TGF-beta(2). Intracranial xenografts of U87MG glioma cells transfected with the dominant-negative ezrins in athymic mice grow to significantly smaller volumes, and the median survival of these mice is 50 d compared with 28 d in the control group. These data define a novel pathway for HGF-induced glioma cell migration and invasion, which requires ezrin, changes in the BCL-2/BAX rheostat, and the induction of TGF-beta(2) expression in vitro, and underscore the important role of HGF signaling in vivo.
Figures

References
-
- Beviglia L, Kramer R. HGF induces FAK activation and integrin-mediated adhesion in MTLn3 breast carcinoma cells. Int J Cancer. 1999;83:640–649. - PubMed
-
- Bourguignon LY, Zhu H, Shao L, Chen YW (2000) CD44 interaction with c-Src kinase promotes cortactin-mediated cytoskeleton function and hyaluronic acid (HA)-dependent ovarian tumor cell migration. J Biol Chem: Nov 17, epub ahead of print. - PubMed
-
- Brooks P, Stromblad S, Sanders LC, von Schalscha TL, Aimes RT, Stetler-Stevenson WG, Quigley JP, Cheresh DA. Localization of matrix metalloproteinase MMP-2 to the surface of invasive cells by interaction with integrin αVβ3. Cell. 1996;85:683–693. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials