

Bidirectional dopamine modulation of GABAergic inhibition in prefrontal cortical pyramidal neurons
- PMID: 11331392
- PMCID: PMC6762481
- DOI: 10.1523/JNEUROSCI.21-10-03628.2001
Bidirectional dopamine modulation of GABAergic inhibition in prefrontal cortical pyramidal neurons
Abstract
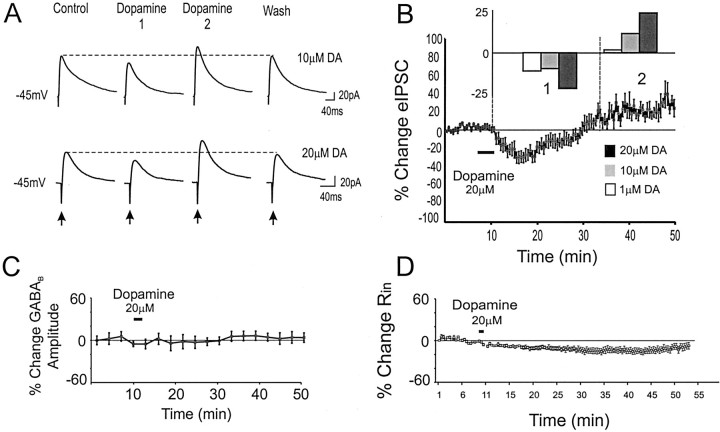
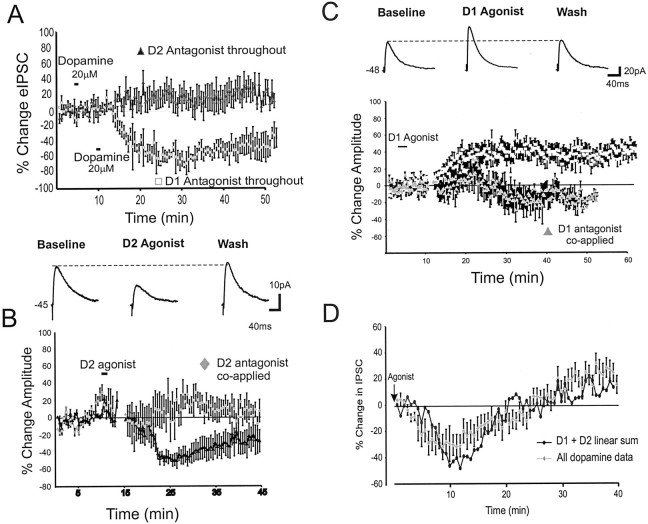
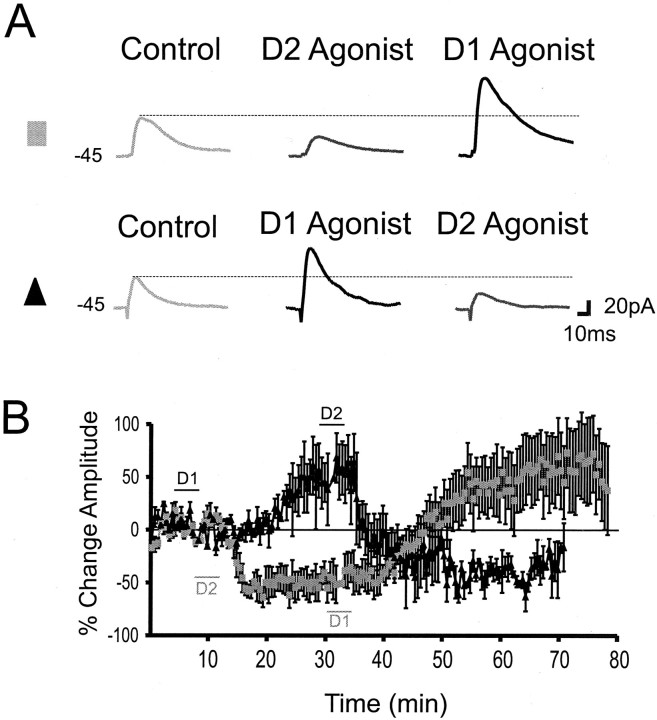
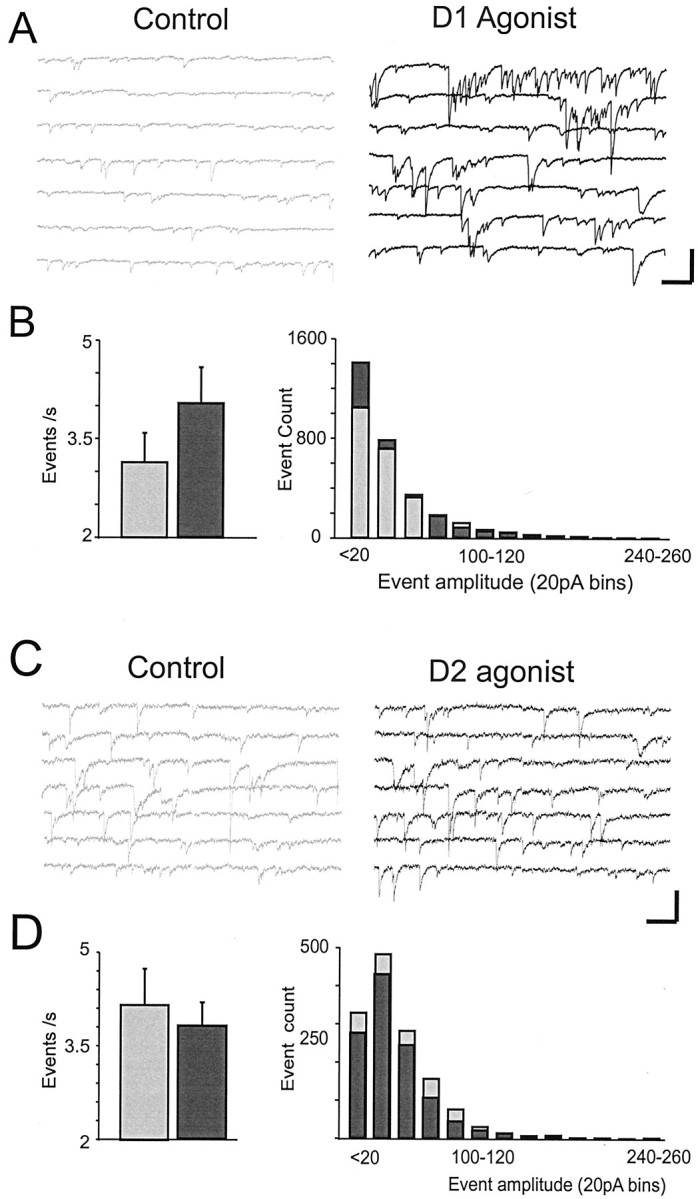
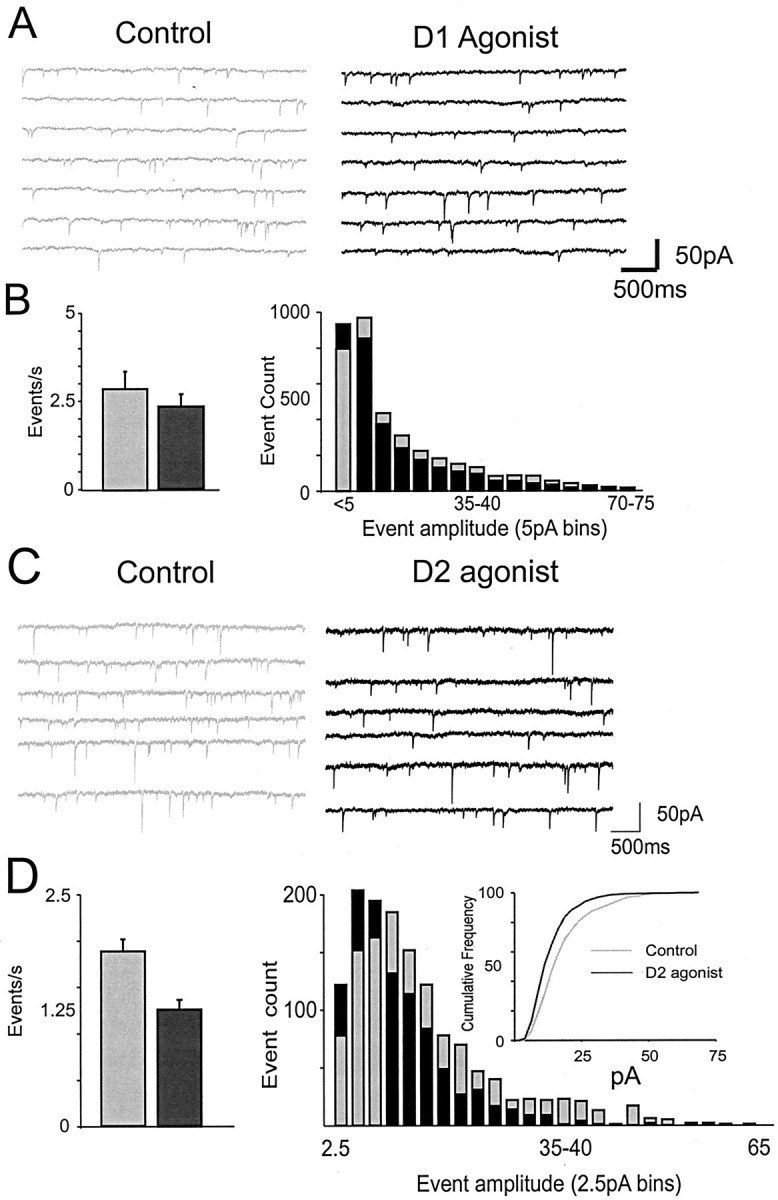
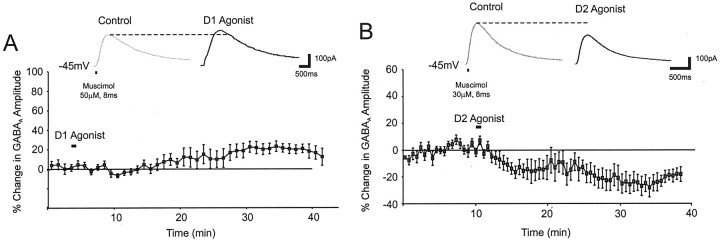
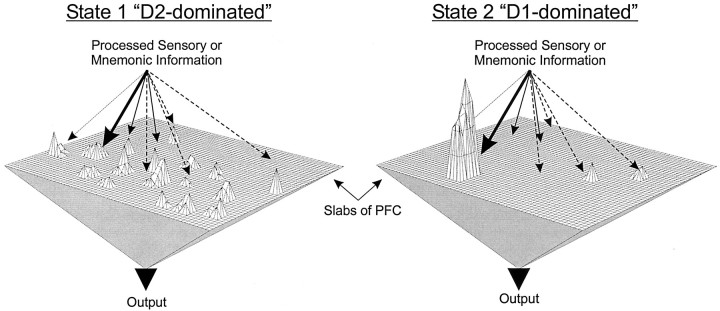
Dopamine regulates the activity of neural networks in the prefrontal cortex that process working memory information, but its precise biophysical actions are poorly understood. The present study characterized the effects of dopamine on GABAergic inputs to prefrontal pyramidal neurons using whole-cell patch-clamp recordings in vitro. In most pyramidal cells, dopamine had a temporally biphasic effect on evoked IPSCs, producing an initial abrupt decrease in amplitude followed by a delayed increase in IPSC amplitude. Using receptor subtype-specific agonists and antagonists, we found that the initial abrupt reduction was D2 receptor-mediated, whereas the late, slower developing enhancement was D1 receptor-mediated. Linearly combining the effects of the two agonists could reproduce the biphasic dopamine effect. Because D1 agonists enhanced spontaneous (sIPSCs) but did not affect miniature (mIPSCs) IPSCs, it appears that D1 agonists caused larger evoked IPSCs by increasing the intrinsic excitability of interneurons and their axons. In contrast, D2 agonists had no effects on sIPSCs but did produce a significant reduction in mIPSCs, suggestive of a decrease in GABA release probability. In addition, D2 agonists reduced the postsynaptic response to a GABA(A) agonist. D1 and D2 receptors therefore regulated GABAergic activity in opposite manners and through different mechanisms in prefrontal cortex (PFC) pyramidal cells. This bidirectional modulation could have important implications for the computational properties of active PFC networks.
Figures
Similar articles
-
Dopamine D4 receptors modulate GABAergic signaling in pyramidal neurons of prefrontal cortex.J Neurosci. 2002 Nov 1;22(21):9185-93. doi: 10.1523/JNEUROSCI.22-21-09185.2002. J Neurosci. 2002. PMID: 12417643 Free PMC article.
-
Dopamine-glutamate interactions controlling prefrontal cortical pyramidal cell excitability involve multiple signaling mechanisms.J Neurosci. 2004 Jun 2;24(22):5131-9. doi: 10.1523/JNEUROSCI.1021-04.2004. J Neurosci. 2004. PMID: 15175382 Free PMC article.
-
Dopamine inhibition of evoked IPSCs in rat prefrontal cortex.J Neurophysiol. 2001 Dec;86(6):2911-8. doi: 10.1152/jn.2001.86.6.2911. J Neurophysiol. 2001. PMID: 11731547
-
Dopamine's Actions in Primate Prefrontal Cortex: Challenges for Treating Cognitive Disorders.Pharmacol Rev. 2015 Jul;67(3):681-96. doi: 10.1124/pr.115.010512. Pharmacol Rev. 2015. PMID: 26106146 Free PMC article. Review.
-
GABA system in the prefrontal cortex involved in psychostimulant addiction.Cereb Cortex. 2024 Aug 1;34(8):bhae319. doi: 10.1093/cercor/bhae319. Cereb Cortex. 2024. PMID: 39098820 Review.
Cited by
-
Effect of dopaminergic D1 receptors on plasticity is dependent of serotoninergic 5-HT1A receptors in L5-pyramidal neurons of the prefrontal cortex.PLoS One. 2015 Mar 16;10(3):e0120286. doi: 10.1371/journal.pone.0120286. eCollection 2015. PLoS One. 2015. PMID: 25775449 Free PMC article.
-
Amphetamine Exerts Dose-Dependent Changes in Prefrontal Cortex Attractor Dynamics during Working Memory.J Neurosci. 2015 Jul 15;35(28):10172-87. doi: 10.1523/JNEUROSCI.2421-14.2015. J Neurosci. 2015. PMID: 26180194 Free PMC article.
-
Dopamine modulates inwardly rectifying potassium currents in medial prefrontal cortex pyramidal neurons.J Neurosci. 2004 Mar 24;24(12):3077-85. doi: 10.1523/JNEUROSCI.4715-03.2004. J Neurosci. 2004. PMID: 15044547 Free PMC article.
-
Impact of Perineuronal Net Removal in the Rat Medial Prefrontal Cortex on Parvalbumin Interneurons After Reinstatement of Cocaine Conditioned Place Preference.Front Cell Neurosci. 2022 Jul 28;16:932391. doi: 10.3389/fncel.2022.932391. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35966203 Free PMC article.
-
Dopamine and corticotropin-releasing factor synergistically alter basolateral amygdala-to-medial prefrontal cortex synaptic transmission: functional switch after chronic cocaine administration.J Neurosci. 2008 Jan 9;28(2):529-42. doi: 10.1523/JNEUROSCI.2666-07.2008. J Neurosci. 2008. PMID: 18184795 Free PMC article.
References
-
- Abekawa T, Ohmori T, Ito K, Koyama T. D1 dopamine receptor activation reduces extracellular glutamate and GABA concentrations in the medial prefrontal cortex. Brain Res. 2000;867:250–254. - PubMed
-
- Benes FM, Vincent SL, Molloy R. Dopamine-immunoreactive axon varicosities form nonrandom contacts with GABA-immunoreactive neurons of rat medial prefrontal cortex. Synapse. 1993;15:285–295. - PubMed
-
- Benes FM, Vincent SL, Molloy R, Khan Y. Increased interaction of dopamine-immunoreactive varicosities with GABA neurons of rat medial prefrontal cortex occurs during the postweanling period. Synapse. 1996;23:237–245. - PubMed
-
- Bourdelais AJ, Deutch AY. The effects of haloperidol and clozapine on extracellular GABA levels in the prefrontal cortex of the rat: an in vivo microdialysis study. Cereb Cortex. 1994;4:69–77. - PubMed
-
- Delgado A, Sierra A, Querejeta E, Valdiosera RF, Aceves J. Inhibitory control of the GABAergic transmission in the rat neostriatum by D2 dopamine receptors. Neuroscience. 2000;95:1043–1048. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous