

SNAREs are concentrated in cholesterol-dependent clusters that define docking and fusion sites for exocytosis
- PMID: 11331586
- PMCID: PMC125434
- DOI: 10.1093/emboj/20.9.2202
SNAREs are concentrated in cholesterol-dependent clusters that define docking and fusion sites for exocytosis
Abstract
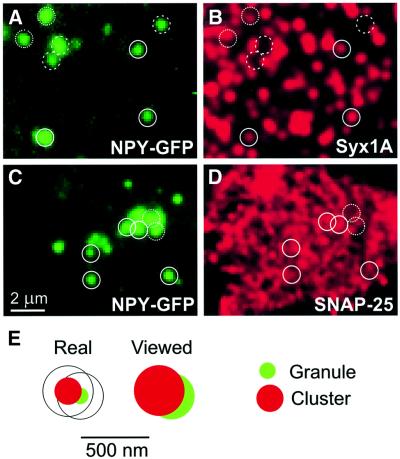
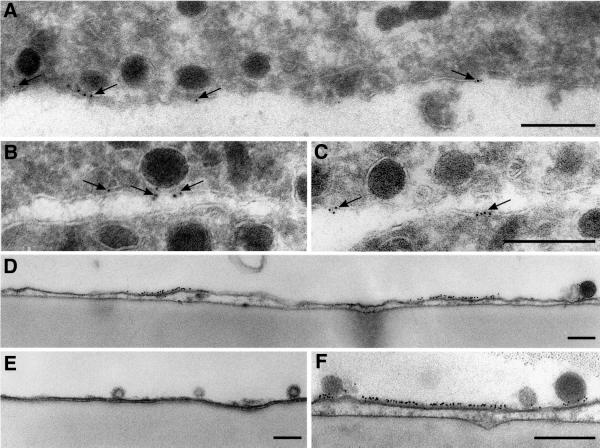
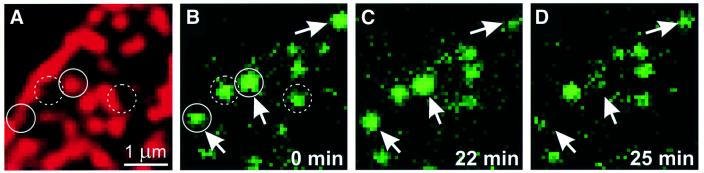
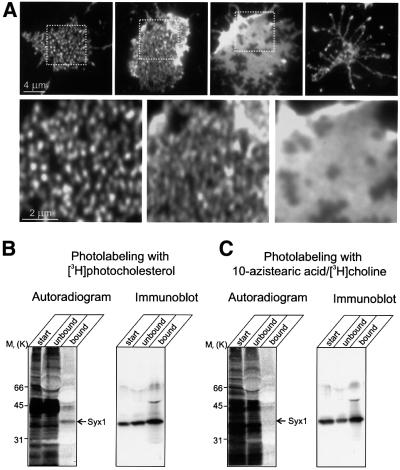
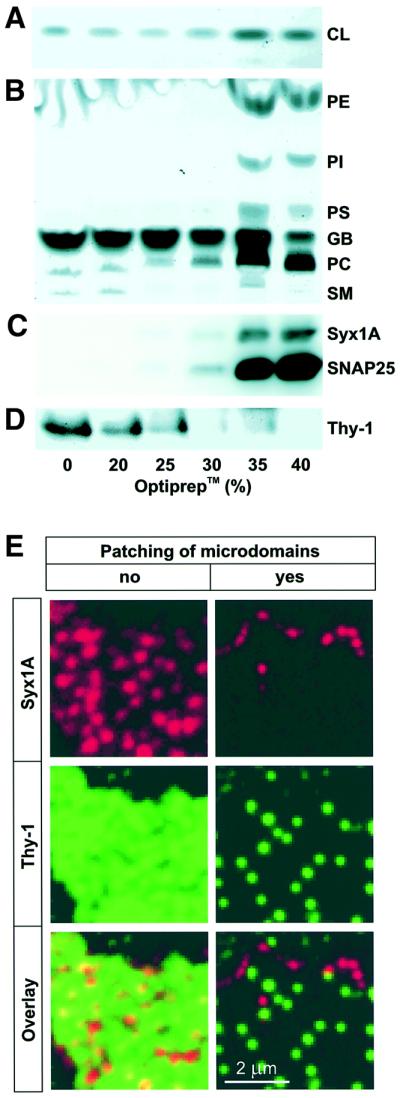
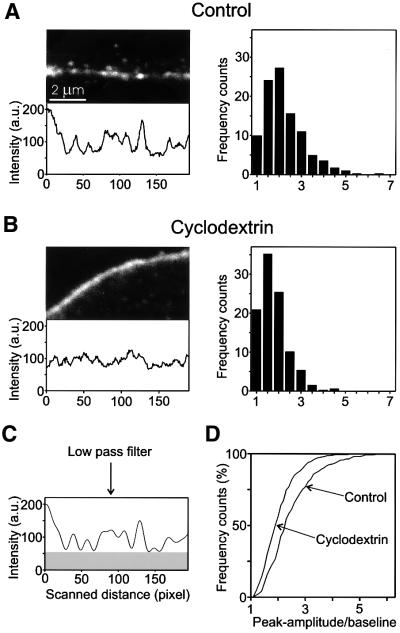
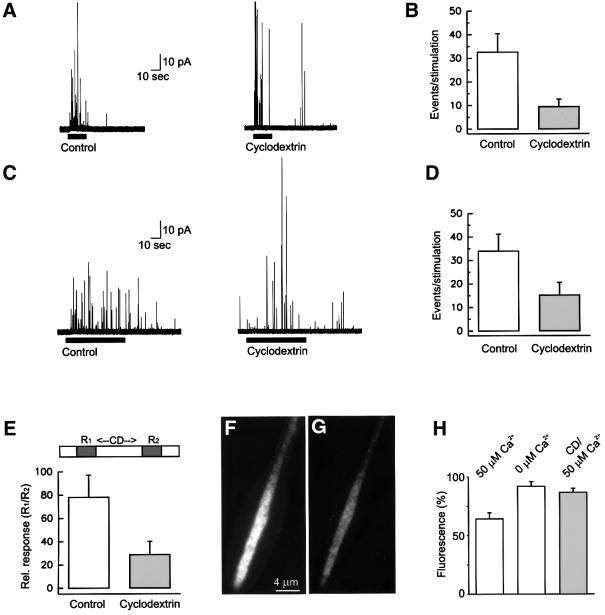
During exocytosis, SNARE proteins of secretory vesicles interact with the corresponding SNARE proteins in the plasmalemma to initiate the fusion reaction. However, it is unknown whether SNAREs are uniformly distributed in the membrane or whether specialized fusion sites exist. Here we report that in the plasmalemma, syntaxins are concentrated in 200 nm large, cholesterol-dependent clusters at which secretory vesicles preferentially dock and fuse. The syntaxin clusters are distinct from cholesterol-dependent membrane rafts since they are Triton X-100-soluble and do not co-patch with raft markers. Synaptosomal-associated protein (SNAP)-25 is also clustered in spots, which partially overlap with syntaxin. Cholesterol depletion causes dispersion of these clusters, which is associated with a strong reduction in the rate of secretion, whereas the characteristics of individual exocytic events are unchanged. This suggests that high local concentrations of SNAREs are required for efficient fusion.
Figures

References
-
- Aguado F., Majo,G., Ruiz-Montasell,B., Canals,J.M., Casanova,A., Marsal,J. and Blasi,J. (1996) Expression of synaptosomal-associated protein SNAP-25 in endocrine anterior pituitary cells. Eur. J. Cell Biol., 69, 351–359. - PubMed
-
- Angleson J.K., Cochilla,A.J., Kilic,G., Nussinovitch,I. and Betz,W.J. (1999) Regulation of dense core release from neuroendocrine cells revealed by imaging single exocytic events. Nature Neurosci., 2, 440–446. - PubMed
-
- Barnstable C.J., Hofstein,R. and Akagawa,K. (1985) A marker of early amacrine cell development in rat retina. Brain Res., 352, 286–290. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
