

TGF-beta-induced repression of CBFA1 by Smad3 decreases cbfa1 and osteocalcin expression and inhibits osteoblast differentiation
- PMID: 11331591
- PMCID: PMC125448
- DOI: 10.1093/emboj/20.9.2254
TGF-beta-induced repression of CBFA1 by Smad3 decreases cbfa1 and osteocalcin expression and inhibits osteoblast differentiation
Abstract
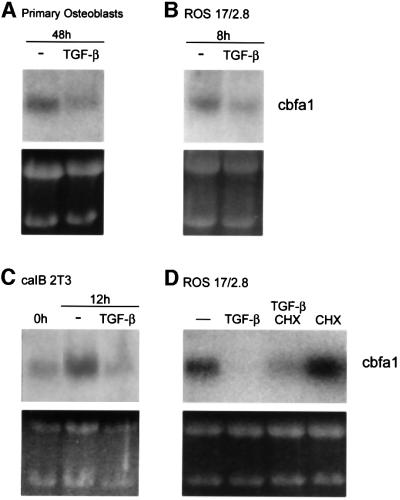
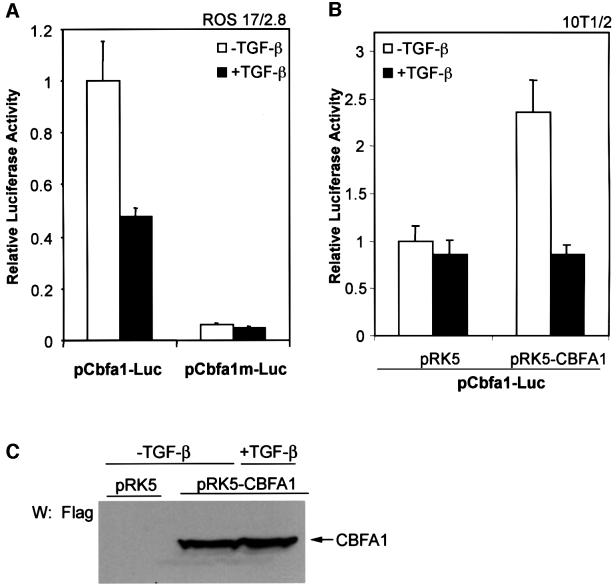
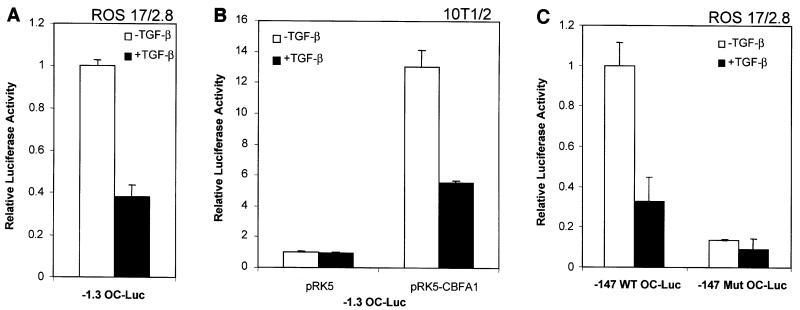
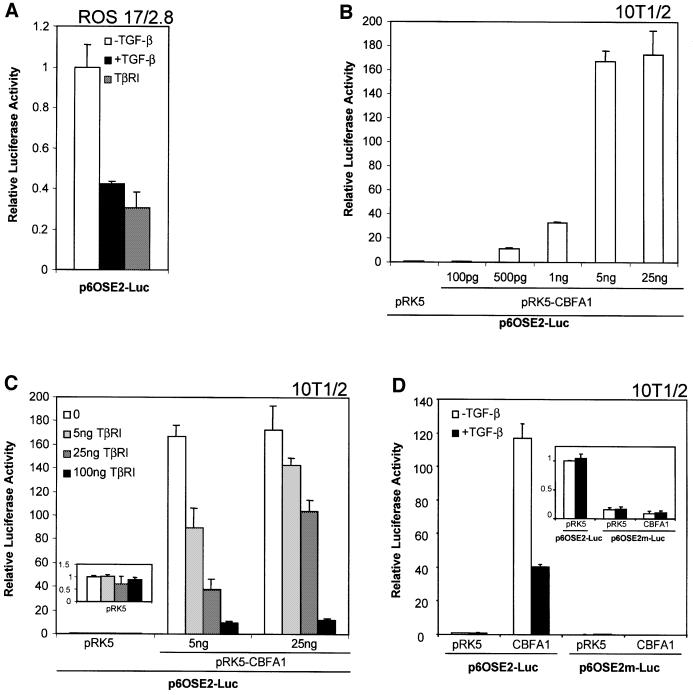
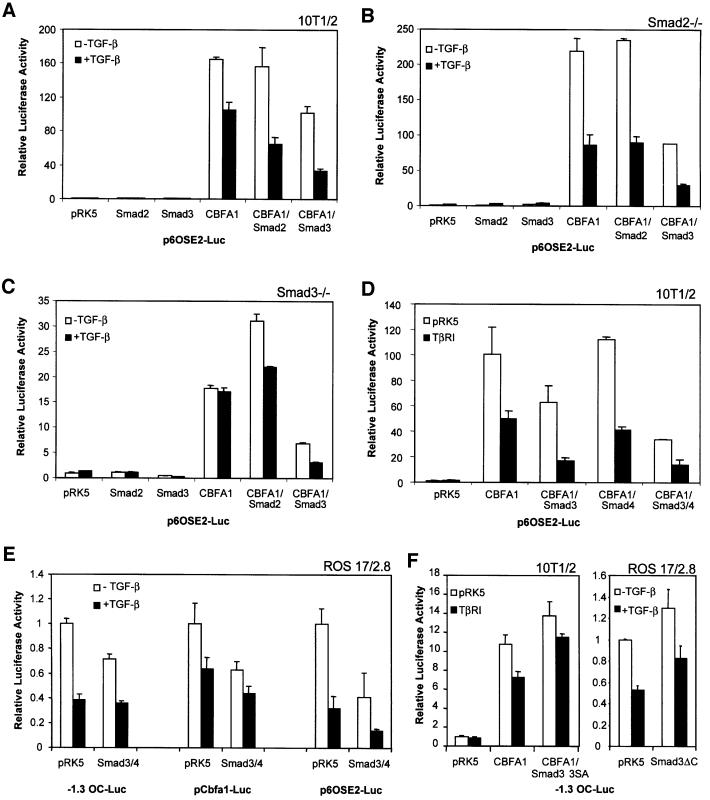
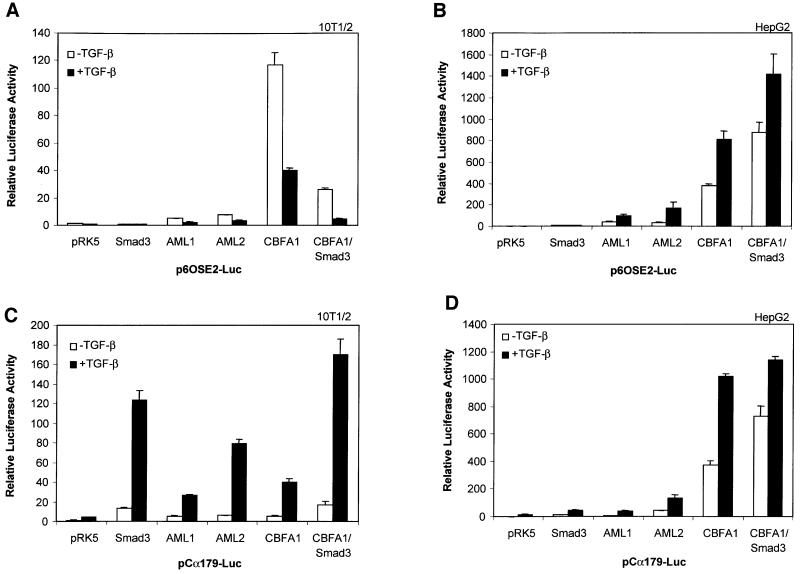
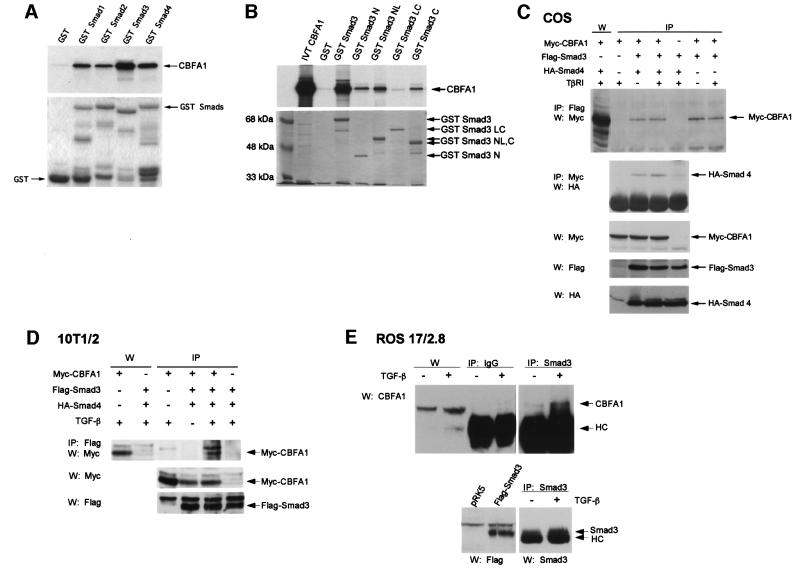
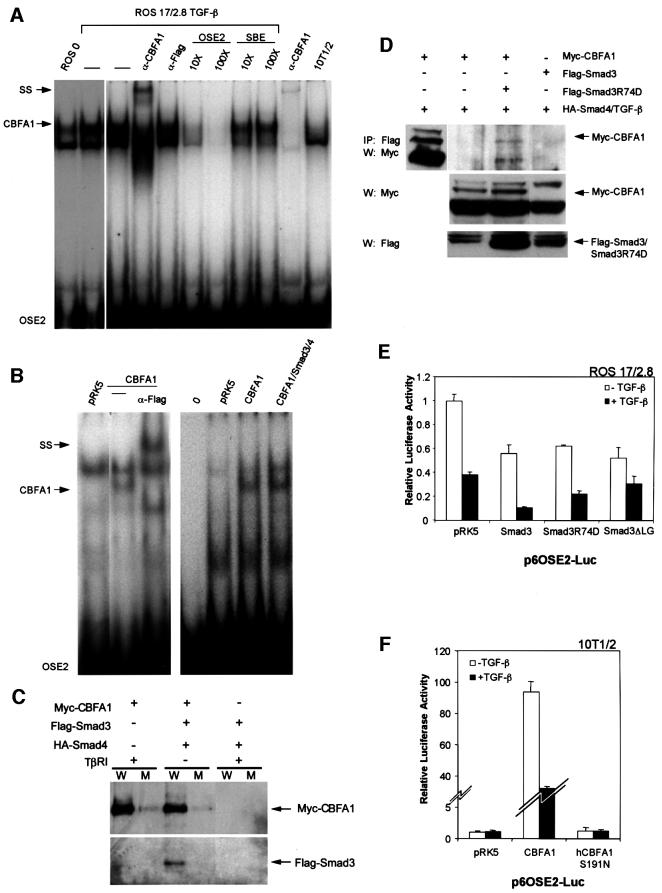
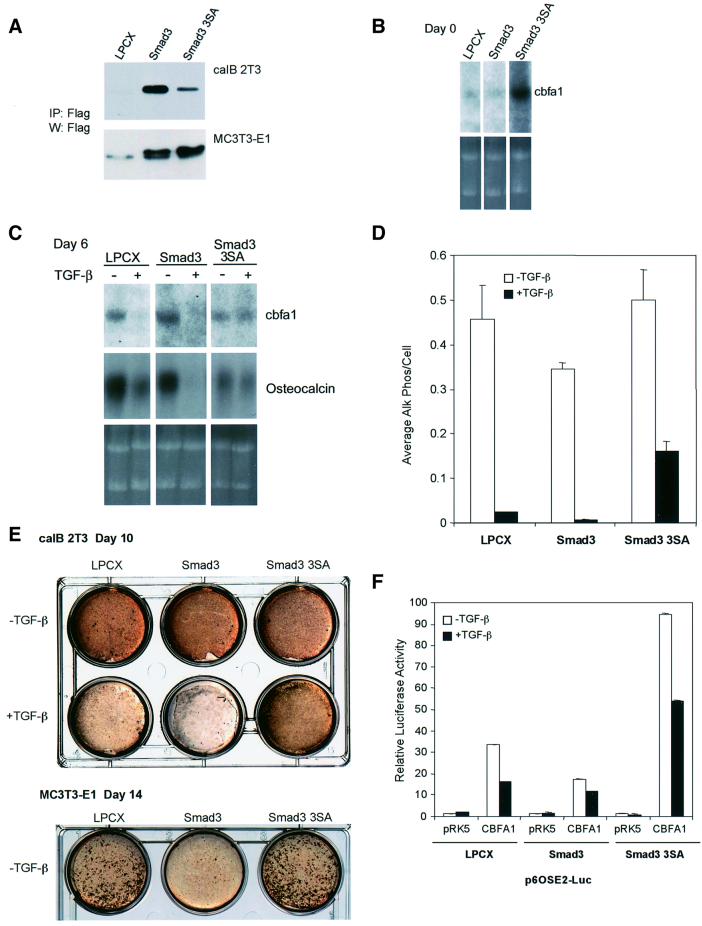
Transforming growth factor-beta (TGF-beta), a secreted factor present at high levels in bone, inhibits osteoblast differentiation in culture; yet, the mechanism of this inhibition remains unclear. We studied the effects of TGF-beta and its effectors, the Smads, on the expression and function of the osteoblast transcription factor CBFA1. TGF-beta inhibited the expression of the cbfa1 and osteocalcin genes, whose expression is controlled by CBFA1 in osteoblast-like cell lines. This inhibition was mediated by Smad3, which interacts physically with CBFA1 and represses its transcriptional activity at the CBFA1-binding OSE2 promoter sequence. The repression of CBFA1 function by Smad3 contrasts with previous observations that Smads function as transcription activators. This repression occurred in mesenchymal but not epithelial cells, and depended on the promoter sequence. Smad3-mediated repression of CBFA1 provides a central regulatory mechanism for the inhibition of osteoblast differentiation by TGF-beta, since it inhibits both cbfa1 transcription and transcriptional activation of osteoblast differentiation genes by CBFA1. Altering Smad3 signaling influenced osteoblast differentiation in the presence or absence of TGF-beta, implicating Smad3/TGF-beta-mediated repression in autocrine regulation of osteoblast differentiation.
Figures
References
-
- Akiyoshi S., Inoue,H., Hanai,J., Kusanagi,K., Nemoto,N., Miyazono,K. and Kawabata,M. (1999) c-Ski acts as a transcriptional co-repressor in transforming growth factor-β signaling through interaction with Smads. J. Biol. Chem., 274, 35269–35277. - PubMed
-
- Alliston T.N. and Derynck,R. (2000) Transforming growth factor-β in skeletal development and maintenance. In Canalis,E. (ed.), Skeletal Growth Factors. Lippincott Williams & Wilkins, Philadelphia, PA, pp. 233–249.
-
- Alliston T.N., Maiyar,A.C., Buse,P., Firestone,G.L. and Richards,J.S. (1997) Follicle stimulating hormone-regulated expression of serum/glucocorticoid-inducible kinase in rat ovarian granulosa cells: a functional role for the Sp1 family in promoter activity. Mol. Endocrinol., 11, 1934–1949. - PubMed
-
- Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, New York, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
