

Infectious entry by amphotropic as well as ecotropic murine leukemia viruses occurs through an endocytic pathway
- PMID: 11333881
- PMCID: PMC114905
- DOI: 10.1128/JVI.75.11.5018-5026.2001
Infectious entry by amphotropic as well as ecotropic murine leukemia viruses occurs through an endocytic pathway
Abstract
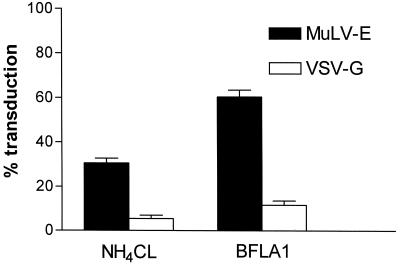
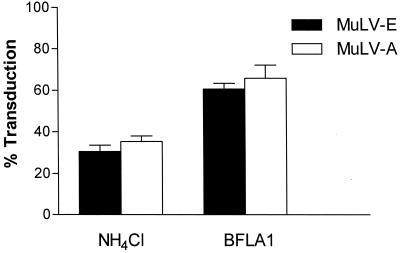
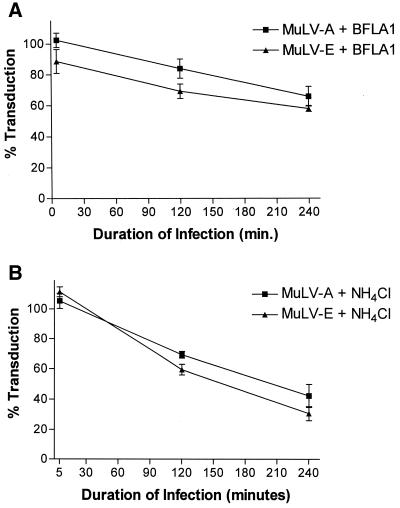
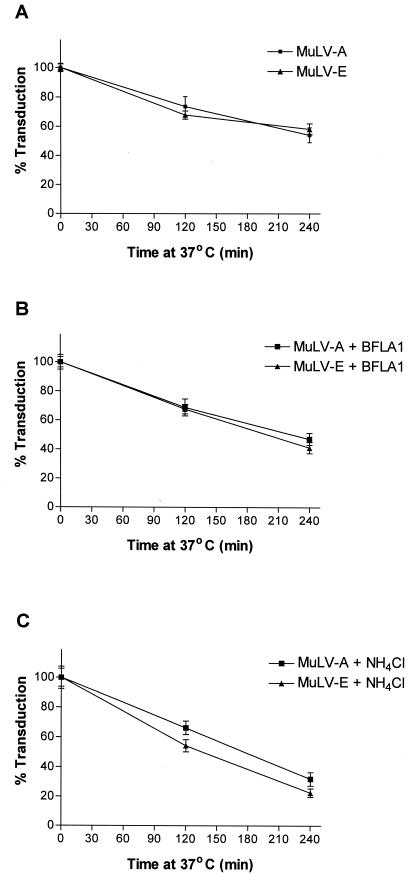
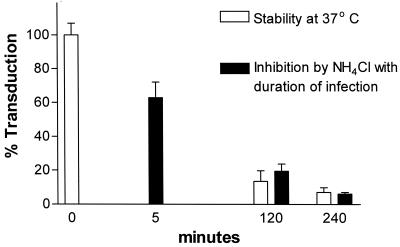
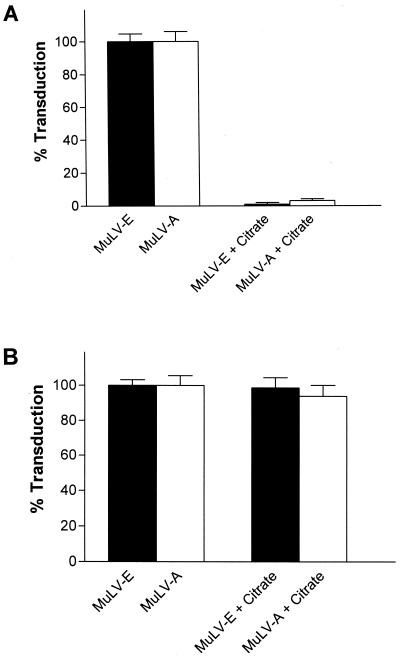
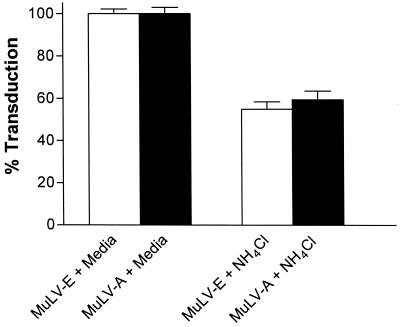
Infectious entry of enveloped viruses is thought to proceed by one of two mechanisms. pH-dependent viruses enter the cells by receptor-mediated endocytosis and are inhibited by transient treatment with agents that prevent acidification of vesicles in the endocytic pathway, while pH-independent viruses are not inhibited by such agents and are thought to enter the cell by direct fusion with the plasma membrane. Nearly all retroviruses, including amphotropic murine leukemia virus (MuLV) and human immunodeficiency virus type 1, are classified as pH independent. However, ecotropic MuLV is considered to be a pH-dependent virus. We have examined the infectious entry of ecotropic and amphotropic MuLVs and found that they were equally inhibited by NH4Cl and bafilomycin A. These agents inhibited both viruses only partially over the course of the experiments. Agents that block the acidification of endocytic vesicles also arrest vesicular trafficking. Thus, partial inhibition of the MuLVs could be the result of virus inactivation during arrest in this pathway. In support of this contention, we found that that the loss of infectivity of the MuLVs during treatment of target cells with the drugs closely corresponded to the loss of activity due to spontaneous inactivation at 37 degrees C in the same period of time. Furthermore, the drugs had no effect on the efficiency of infection under conditions in which the duration of infection was held to a very short period to minimize the effects of spontaneous inactivation. These results indicate that the infectious processes of both ecotropic and amphotropic MuLVs were arrested rather than aborted by transient treatment of the cells with the drugs. We also found that infectious viruses were efficiently internalized during treatment. This indicated that the arrest occurred in an intracellular compartment and that the infectious process of both the amphotropic and ecotropic MuLVs very likely involved endocytosis. An important aspect of this study pertains to the interpretation of experiments in which agents that block endocytic acidification inhibit infectivity. As we have found with the MuLVs, inhibition of infectivity may be secondary to the block of endocytic acidification. While this strongly suggests the involvement of an endocytic pathway, it does not necessarily indicate a requirement for an acidic compartment during the infectious process. Likewise, a lack of inhibition during transient treatment with the drugs would not preclude an endocytic pathway for viruses that are stable during the course of the treatment.
Figures
References
-
- Andersen K B, Nexo B A. Entry of murine retrovirus into mouse fibroblasts. Virology. 1983;125:85–98. - PubMed
-
- Andreadis S, Palsson B O. Coupled effects of polybrene and calf serum on the efficiency of retroviral transduction and the stability of retroviral vectors. Hum Gene Ther. 1997;8:285–291. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
