

Requirement of the nicotinic acetylcholine receptor beta 2 subunit for the anatomical and functional development of the visual system
- PMID: 11344259
- PMCID: PMC33489
- DOI: 10.1073/pnas.101120998
Requirement of the nicotinic acetylcholine receptor beta 2 subunit for the anatomical and functional development of the visual system
Abstract
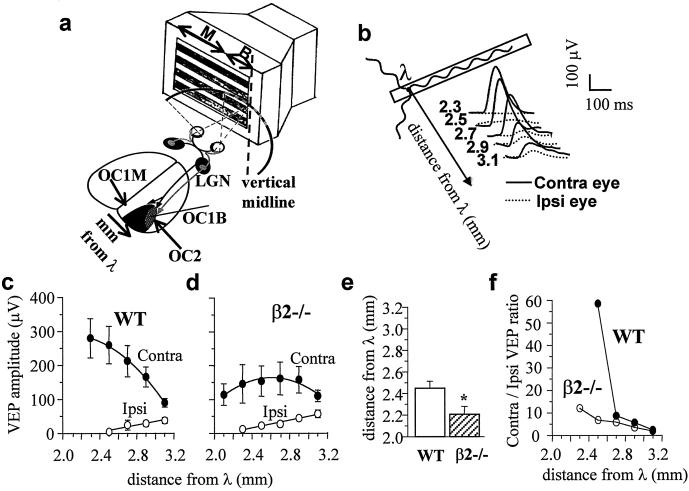
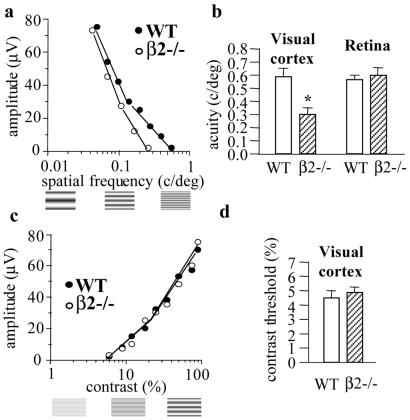
In the mammalian visual system the formation of eye-specific layers at the thalamic level depends on retinal waves of spontaneous activity, which rely on nicotinic acetylcholine receptor activation. We found that in mutant mice lacking the beta2 subunit of the neuronal nicotinic receptor, but not in mice lacking the alpha4 subunit, retinofugal projections do not segregate into eye-specific areas, both in the dorso-lateral geniculate nucleus and in the superior colliculus. Moreover, beta2-/- mice show an expansion of the binocular subfield of the primary visual cortex and a decrease in visual acuity at the cortical level but not in the retina. We conclude that the beta2 subunit of the nicotinic acetylcholine receptor is necessary for the anatomical and functional development of the visual system.
Figures


References
-
- Shatz C J, Stryker M P. Science. 1988;242:87–89. - PubMed
-
- Sretavan D W, Shatz C J, Stryker M P. Nature (London) 1988;336:468–471. - PubMed
-
- Hahm J-O, Langdon R B, Sur M. Nature (London) 1991;351:568–570. - PubMed
-
- Katz L C, Shatz C J. Science. 1996;274:1133–1138. - PubMed
-
- Galli L, Maffei L. Science. 1988;242:90–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
