

Cerebral energetics and the glycogen shunt: neurochemical basis of functional imaging
- PMID: 11344262
- PMCID: PMC33483
- DOI: 10.1073/pnas.101129298
Cerebral energetics and the glycogen shunt: neurochemical basis of functional imaging
Abstract
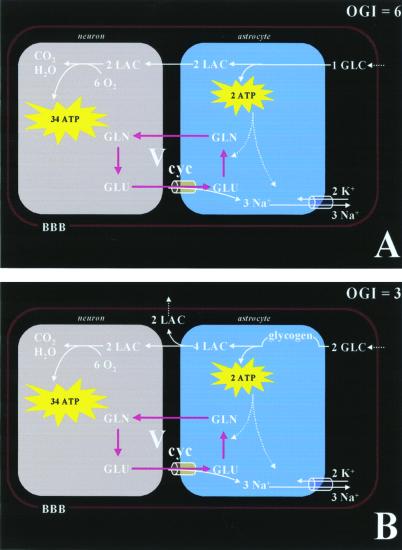
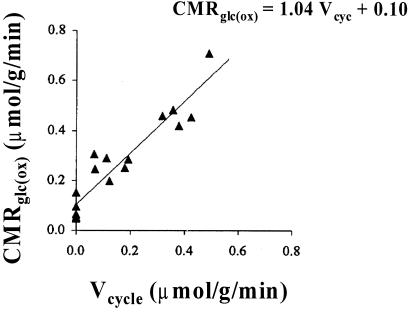
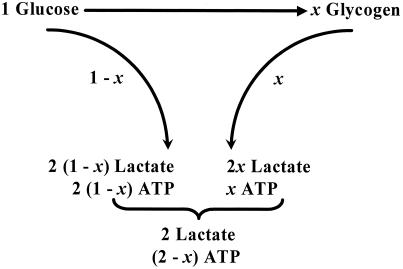
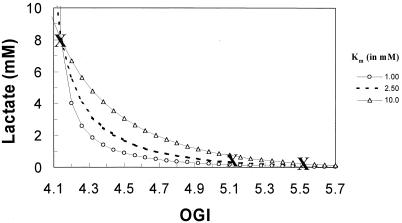
Positron-emission tomography and functional MRS imaging signals can be analyzed to derive neurophysiological values of cerebral blood flow or volume and cerebral metabolic consumption rates of glucose (CMR(Glc)) or oxygen (CMR(O(2))). Under basal physiological conditions in the adult mammalian brain, glucose oxidation is nearly complete so that the oxygen-to-glucose index (OGI), given by the ratio of CMR(O(2))/CMR(Glc), is close to the stoichiometric value of 6. However, a survey of functional imaging data suggests that the OGI is activity dependent, moving further below the oxidative value of 6 as activity is increased. Brain lactate concentrations also increase with stimulation. These results had led to the concept that brain activation is supported by anaerobic glucose metabolism, which was inconsistent with basal glucose oxidation. These differences are resolved here by a proposed model of glucose energetics, in which a fraction of glucose is cycled through the cerebral glycogen pool, a fraction that increases with degree of brain activation. The "glycogen shunt," although energetically less efficient than glycolysis, is followed because of its ability to supply glial energy in milliseconds for rapid neurotransmitter clearance, as a consequence of which OGI is lowered and lactate is increased. The value of OGI observed is consistent with passive lactate efflux, driven by the observed lactate concentration, for the few experiments with complete data. Although the OGI changes during activation, the energies required per neurotransmitter release (neuronal) and clearance (glial) are constant over a wide range of brain activity.
Figures

References
-
- Siesjo B. Brain Energy Metabolism. New York: Wiley; 1978.
-
- Kety S S. In: The Metabolism of the Nervous System. Richter D, editor. London: Pergamon; 1957. pp. 221–237.
-
- Sokoloff L. In: Brain Work and Mental Activity. Lassen N A, Ingvar D H, Raichle M E, Friberg L, editors. Copenhagen: Munksgaard; 1991. pp. 52–64.
-
- Fox P T, Raichle M E, Mintun M A, Dence C. Science. 1988;241:462–464. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
