

Disruption of Plasmodium falciparum chitinase markedly impairs parasite invasion of mosquito midgut
- PMID: 11349075
- PMCID: PMC98468
- DOI: 10.1128/IAI.69.6.4048-4054.2001
Disruption of Plasmodium falciparum chitinase markedly impairs parasite invasion of mosquito midgut
Abstract
To initiate invasion of the mosquito midgut, Plasmodium ookinetes secrete chitinolytic activity to penetrate the peritrophic matrix surrounding the blood meal. While ookinetes of the avian malaria parasite Plasmodium gallinaceum appear to secrete products of two chitinase genes, to date only one chitinase gene, PfCHT1, has been identified in the nearly completed Plasmodium falciparum strain 3D7 genome database. To test the hypothesis that the single identified chitinase of P. falciparum is necessary for ookinete invasion, the PfCHT1 gene was disrupted 39 bp upstream of the stop codon. PfCHT1-disrupted parasites had normal gametocytogenesis, exflagellation, and ookinete formation but were markedly impaired in their ability to form oocysts in Anopheles freeborni midguts. Confocal microscopy demonstrated that the truncated PfCHT1 protein was present in mutant ookinetes but that the concentration of mutant PfCHT1 within the apical end of the ookinetes was substantially reduced. These data suggest that full-length PfCHT1 is essential for intracellular trafficking and secretion and that the PfCHT1 gene product is necessary for ookinetes to invade the mosquito midgut.
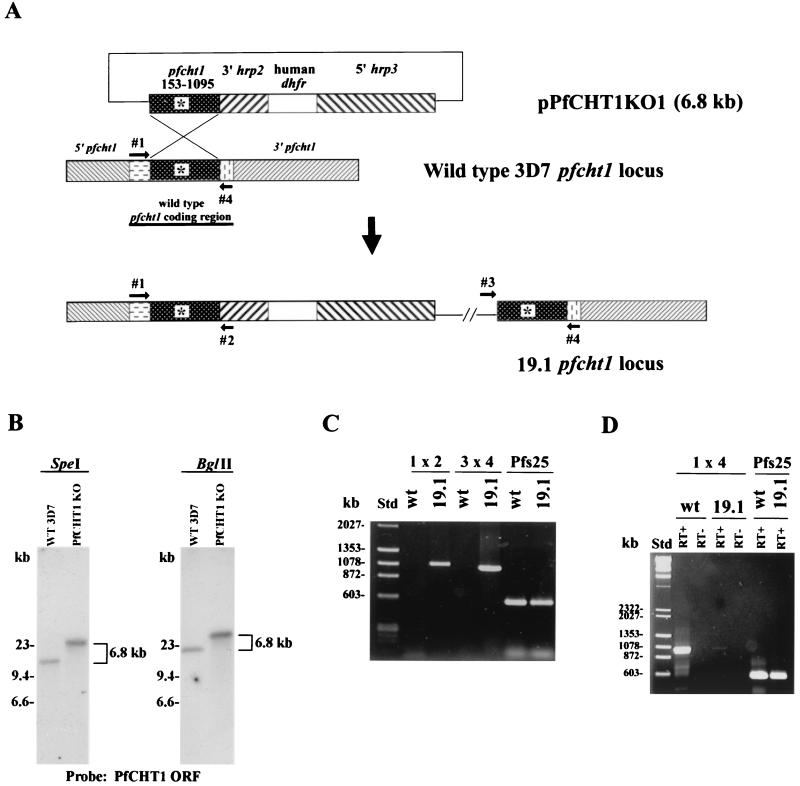
Figures

Similar articles
-
A Hetero-Multimeric Chitinase-Containing Plasmodium falciparum and Plasmodium gallinaceum Ookinete-Secreted Protein Complex Involved in Mosquito Midgut Invasion.Front Cell Infect Microbiol. 2021 Jan 8;10:615343. doi: 10.3389/fcimb.2020.615343. eCollection 2020. Front Cell Infect Microbiol. 2021. PMID: 33489941 Free PMC article.
-
The chitinase PfCHT1 from the human malaria parasite Plasmodium falciparum lacks proenzyme and chitin-binding domains and displays unique substrate preferences.Proc Natl Acad Sci U S A. 1999 Nov 23;96(24):14061-6. doi: 10.1073/pnas.96.24.14061. Proc Natl Acad Sci U S A. 1999. PMID: 10570198 Free PMC article.
-
Knockout of the rodent malaria parasite chitinase pbCHT1 reduces infectivity to mosquitoes.Infect Immun. 2001 Jun;69(6):4041-7. doi: 10.1128/IAI.69.6.4041-4047.2001. Infect Immun. 2001. PMID: 11349074 Free PMC article.
-
Interactions of human malaria parasites, Plasmodium vivax and P.falciparum, with the midgut of Anopheles mosquitoes.Med Vet Entomol. 1997 Jul;11(3):290-6. doi: 10.1111/j.1365-2915.1997.tb00409.x. Med Vet Entomol. 1997. PMID: 9330262 Review.
-
Do malaria ookinete surface proteins P25 and P28 mediate parasite entry into mosquito midgut epithelial cells?Malar J. 2005 Feb 25;4:15. doi: 10.1186/1475-2875-4-15. Malar J. 2005. PMID: 15733320 Free PMC article. Review.
Cited by
-
Identification of novel Plasmodium gallinaceum zygote- and ookinete-expressed proteins as targets for blocking malaria transmission.Infect Immun. 2002 Jan;70(1):102-6. doi: 10.1128/IAI.70.1.102-106.2002. Infect Immun. 2002. PMID: 11748169 Free PMC article.
-
Synthesis, antiasthmatic, and insecticidal/antifungal activities of allosamidins.J Enzyme Inhib Med Chem. 2019 Dec;34(1):1226-1232. doi: 10.1080/14756366.2019.1623208. J Enzyme Inhib Med Chem. 2019. PMID: 31307248 Free PMC article. Review.
-
The Plasmodium bottleneck: malaria parasite losses in the mosquito vector.Mem Inst Oswaldo Cruz. 2014 Aug;109(5):644-61. doi: 10.1590/0074-0276130597. Mem Inst Oswaldo Cruz. 2014. PMID: 25185005 Free PMC article.
-
A subpellicular microtubule dynein transport machinery regulates ookinete morphogenesis for mosquito transmission of Plasmodium yoelii.Nat Commun. 2024 Oct 4;15(1):8590. doi: 10.1038/s41467-024-52970-7. Nat Commun. 2024. PMID: 39366980 Free PMC article.
-
Tick gut barriers impacting tick-microbe interactions and pathogen persistence.Mol Microbiol. 2021 Nov;116(5):1241-1248. doi: 10.1111/mmi.14822. Epub 2021 Oct 6. Mol Microbiol. 2021. PMID: 34570926 Free PMC article. Review.
References
-
- Babiker H A, Abdel-Wahab A, Ahmed S, Suleiman S, Ranford-Cartwright L, Carter R, Walliker D. Detection of low level Plasmodium falciparum gametocytes using reverse transcriptase polymerase chain reaction. Mol Biochem Parasitol. 1999;99:143–148. - PubMed
-
- Carlton J M, Galinski M R, Barnwell J W, Dame J B. Karyotype and synteny among the chromosomes of all four species of human malaria parasite. Mol Biochem Parasitol. 1999;101:23–32. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
