

doi: 10.1128/IAI.69.6.4159-4163.2001.
Influence of cultivation media on genetic regulatory patterns in Borrelia burgdorferi
Affiliations
- PMID: 11349092
- PMCID: PMC98485
- DOI: 10.1128/IAI.69.6.4159-4163.2001
Item in Clipboard
Influence of cultivation media on genetic regulatory patterns in Borrelia burgdorferi
Infect Immun.
2001 Jun.
Abstract
Barbour-Stoenner-Kelly II (BSKII) medium and BSKH medium both are routinely used for the cultivation of Borrelia burgdorferi. However, heretofore there have been no studies to compare how these two media affect gene expression patterns in virulent B. burgdorferi. In the present study, we found that some B. burgdorferi strain 297 genes (e.g., ospA, mlp-7A, mlp-8, p22, and lp6.6) that typically are regulated by temperature or pH displayed their predicted pattern of expression when B. burgdorferi was cultivated in BSKH medium; this was not true when spirochetes were cultivated in conventional BSKII medium. The results suggest that BSKH medium is superior to BSKII medium for gene expression studies with B. burgdorferi.
Figures
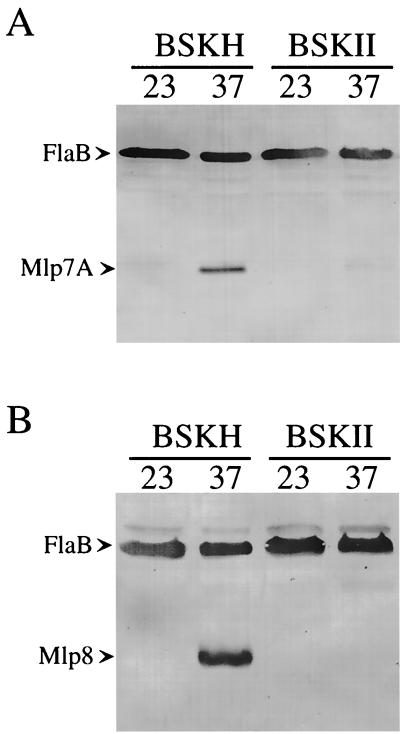
Influence of BSKH or BSKII medium on the levels of Mlp-7A or Mlp-8 expressed in B. burgdorferi 297 cultivated at either 23 or 37°C. B. burgdorferi adapted at 23°C was inoculated at a final concentration of 1 × 103 spirochetes per ml, and 23 or 37°C cultures were harvested at a density of 5 × 107 cells per ml (late logarithmic phase). Protein from 5 × 107 spirochetes was loaded in each sodium dodecyl sulfate-polyacrylamide gel lane and subjected to immunoblot assay. The antibodies used to detect FlaB and Mlp-8 were described previously (27). Rat monospecific polyclonal antiserum to an epitopic region of Mlp-7A was generated using a method analogous to that used to create antiserum against Mlp-8 (27). (A) Immunoblot using a mixture of antibodies against FlaB and Mlp-7A. (B) Immunoblot using a mixture of antibodies against FlaB and Mlp-8. Immunoblotting for FlaB was performed to confirm that the numbers of spirochetes loaded in the gel lanes were substantially equivalent.
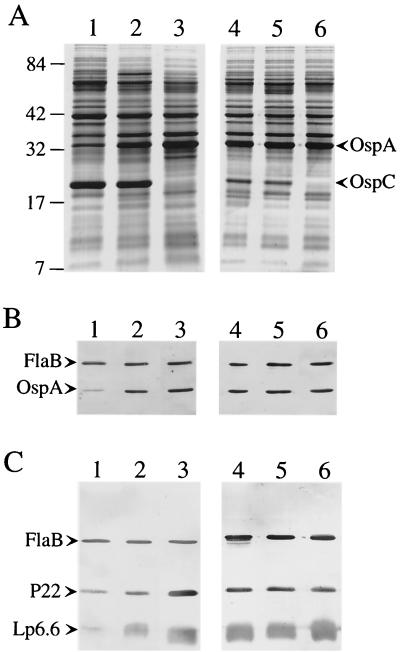
Influence of BSKH (lanes 1 to 3) or BSKII (lanes 4 to 6) medium on the levels of OspA, P22, and Lp6.6 expressed in B. burgdorferi 297 cultivated under various pH conditions. Media were preadjusted to either pH 6.8 (lanes 1 and 4), 7.5 (lanes 2 and 5), or 8.0 (lanes 3 and 6) as reported elsewhere (26). B. burgdorferi 297 was then inoculated at a final concentration of 1 × 103 spirochetes per ml, incubated at 37°C, and harvested at a density of 5 × 107 spirochetes per ml. (A) Sodium dodecyl sulfate-polyacrylamide gel stained with Coomassie brilliant blue; each gel lane contained protein from 5 × 107 spirochetes. Protein molecular mass standards (in kilodaltons) are indicated at the left. Polypeptides corresponding to OspA and OspC are labeled at the right. (B) Immunoblot of samples used in panel A, except that gel lanes contained protein from 5 × 106 spirochetes (in order to visualize the relative differences in OspA levels); the antibodies used to detect the respective antigens were described previously (26). (C) Same as panel B, except that protein from 107 spirochetes was loaded in each gel lane. Immunoblotting for FlaB (panels B and C) was performed to confirm that the numbers of spirochetes loaded in the gel lanes were substantially equivalent.
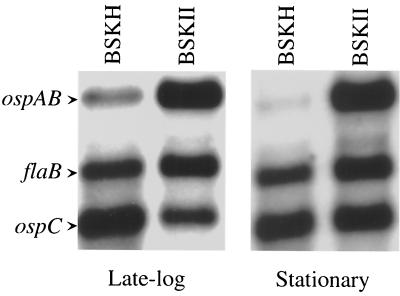
Northern blot analysis of the expression of ospAB, flaB, and ospC in B. burgdorferi during the late logarithmic (5 × 107 spirochetes/ml) and stationary (1 × 108 spirochetes/ml) phases of growth. RNA from B. burgdorferi 297 cultivated at each stage of growth in either BSKH or BSKII medium at 37°C (pH 6.8) was hybridized with probes specific for either ospAB, flaB, or ospC. The methods for mRNA isolation and Northern blot assays have been described previously (21, 27). Three micrograms of total RNA was loaded in each gel lane. The three hybridization probes were mixed together in the Northern blot assays. The bands corresponding to each transcript are labeled at the left. Assessment of the level of flaB mRNA was used as an internal control to indicate equivalent RNA loading among the various gel lanes.
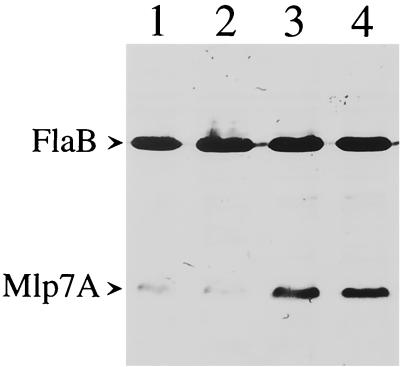
Influence of BSA and rabbit serum in BSKII medium on the level of Mlp-7A expressed by B. burgdorferi. B. burgdorferi 297 adapted at 23°C was inoculated at a final concentration of 1 × 103 spirochetes per ml, incubated at 37°C, and harvested at a final density of 5 × 107 cells per ml (late logarithmic phase). Protein from 5 × 107 spirochetes was loaded in each gel lane, and the samples were then subjected to immunoblot assay. The antibodies used to detect each antigen were as for Fig. 1A. Lane 1, spirochetes cultivated in BSKII medium formulated with conventional (nonprescreened) BSA and rabbit serum. Lane 2, spirochetes grown in BSKII medium containing the same nonprescreened BSA but the rabbit serum used to make commercial BSKH medium. Lane 3, borreliae cultivated in BSKII medium formulated with conventional rabbit serum but the BSA used in the commercial preparation of BSKH medium. Lane 4, spirochetes grown in BSKII medium containing both the quality-controlled rabbit serum and quality-controlled BSA, thereby making it substantially equivalent to commercial BSKH medium. Immunoblotting for FlaB was performed to confirm that the numbers of spirochetes were essentially equal within the gel lanes.
References
-
- Akins D R, Porcella S F, Popova T G, Shevchenko D, Li M, Norgard M V, Radolf J D. Evidence for in vivo but not in vitro expression of a Borrelia burgdorferi outer surface protein F (OspF) homologue. Mol Microbiol. 1995;18:507–520. - PubMed
-
- Alban P S, Johnson P W, Nelson D R. Serum-starvation-induced changes in protein synthesis and morphology of Borrelia burgdorferi. Microbiology. 2000;146:119–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
