

Evolutionarily divergent electron donor proteins interact with P450MT2 through the same helical domain but different contact points
- PMID: 11350928
- PMCID: PMC125462
- DOI: 10.1093/emboj/20.10.2394
Evolutionarily divergent electron donor proteins interact with P450MT2 through the same helical domain but different contact points
Abstract
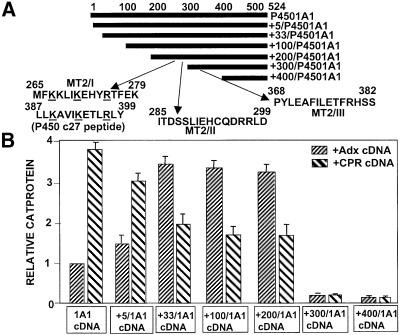
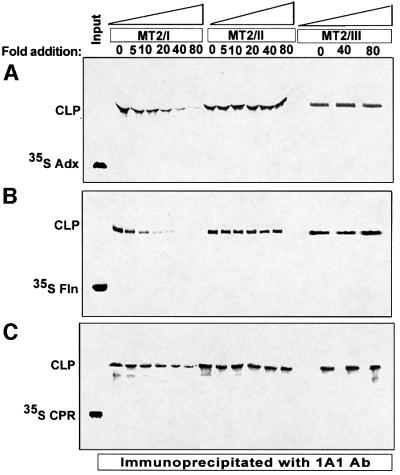
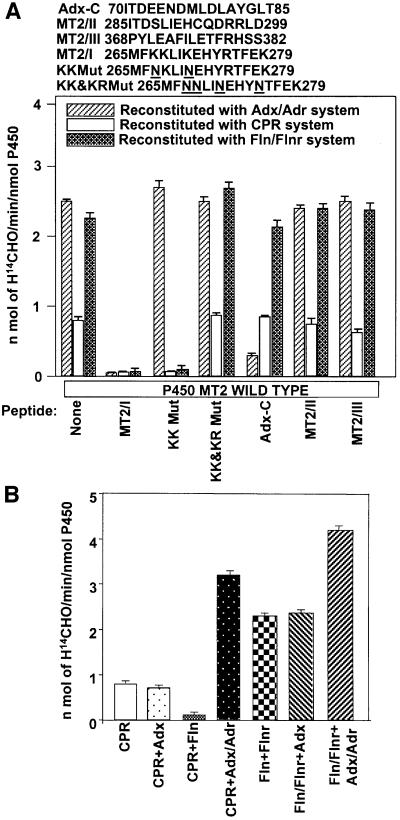
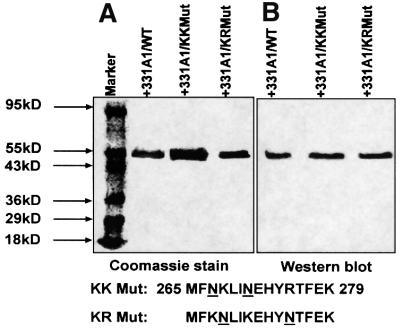
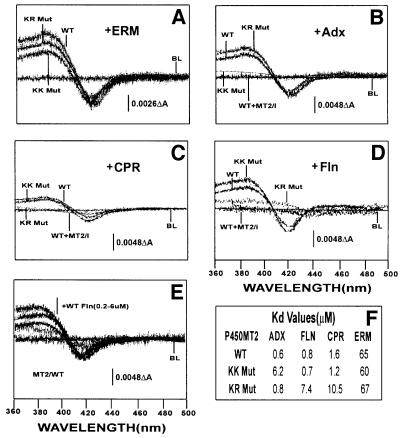
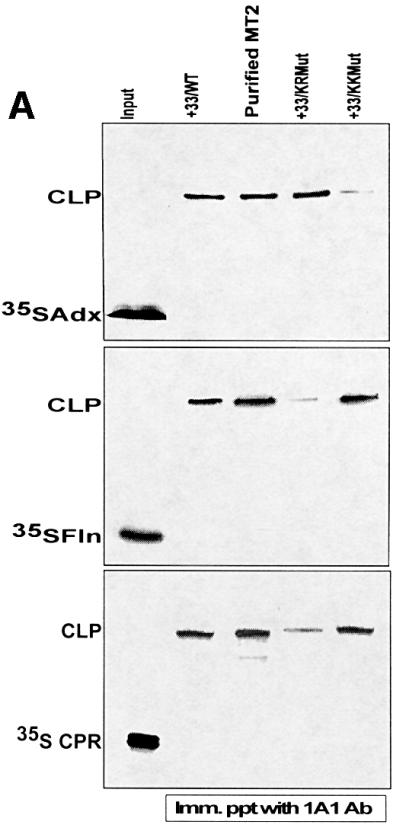
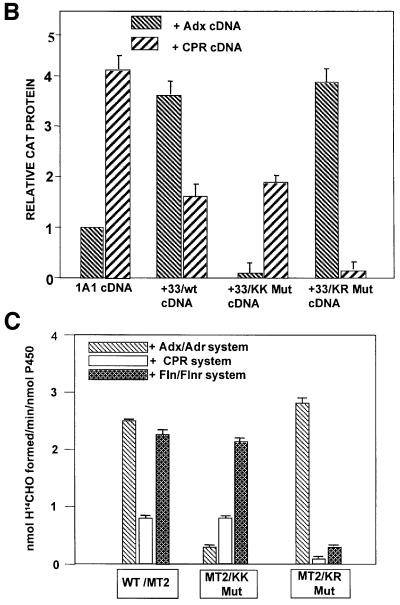
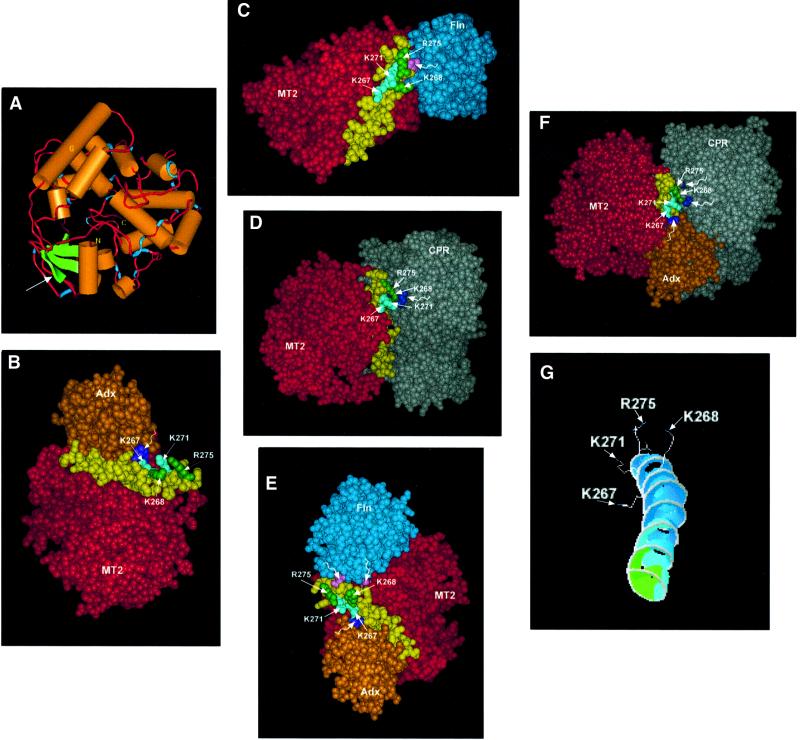
We have investigated the sites of N-terminally truncated cytochrome P4501A1 targeted to mitochondria (P450MT2) which interact with adrenodoxin (Adx), cytochrome P450 reductase (CPR) and bacterial flavodoxin (Fln). The binding site was mapped by a combination of in vitro mutagenesis, in vivo screening with a mammalian two-hybrid system, spectral analysis, reconstitution of enzyme activity and homology-based structural modeling. Our results show that part of an aqueous accessible helix (putative helix G, residues 264-279) interacts with all three electron donor proteins. Mutational studies revealed that Lys267 and Lys271 are crucial for binding to Adx, while Lys268 and Arg275 are important for binding to CPR and FLN: Additive effects of different electron donor proteins on enzyme activity and models on protein docking show that Adx and CPR bind in a non-overlapping manner to the same helical domain in P450MT2 at different angular orientations, while CPR and Fln compete for the same binding site. We demonstrate that evolutionarily divergent electron donor proteins interact with the same domain but subtly different contact points of P450MT2.
Figures
References
-
- Anandatheerthavarada H.K., Addya,S., Dwivedi,R.S., Biswas,G., Mullick,J. and Avadhani,N.G. (1997) Localization of multiple forms of inducible cytochromes P450 in rat liver mitochondria: immunological characteristics and patterns of xenobiotic substrate metabolism. Arch. Biochem. Biophys., 339, 136–150. - PubMed
-
- Anandatheerthavarada H.K., Addya,S., Mullick,J. and Avadhani,N.G. (1998) Interaction of adrenodoxin with P450 1A1 and its truncated form P450MT2 through different domains: differential modulation of enzyme activities:. Biochemistry, 37, 1150–1160. - PubMed
-
- Anandatheerthavarada H.K., Vijayasarathy,C., Bhagwat,S.V., Biswas, G., Mullick,J. and Avadhani,N.G. (1999b) Physiological role of the N-terminal processed P4501A1 targeted to mitochondria in erythromycin metabolism and reversal of erythromycin-mediated inhibition of mitochondrial protein synthesis. J. Biol. Chem., 274, 6617–6625. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
