

State-dependent modulation of sensory feedback
- PMID: 11351007
- PMCID: PMC2278613
- DOI: 10.1111/j.1469-7793.2001.0005b.x
State-dependent modulation of sensory feedback
Abstract
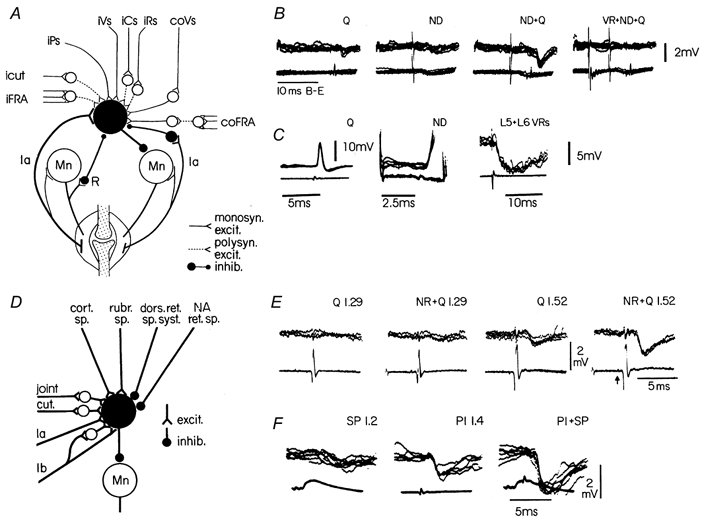
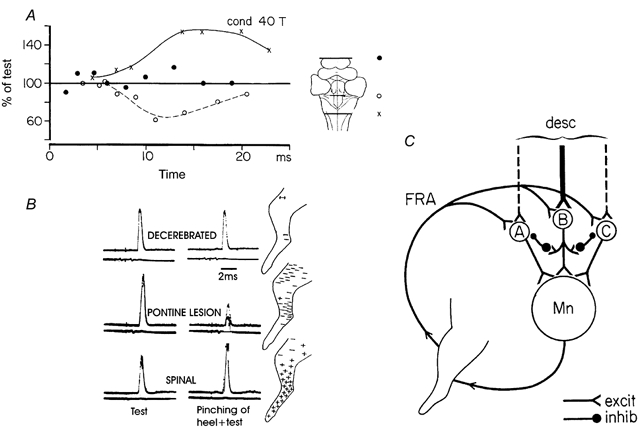
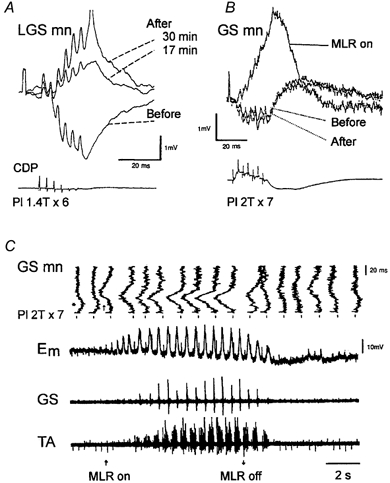
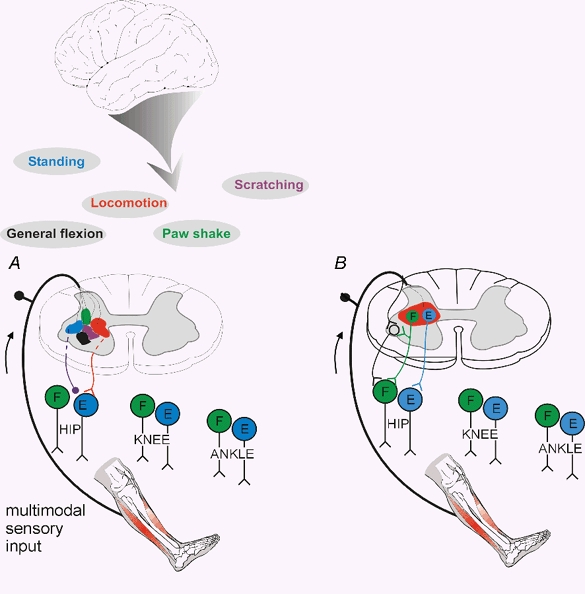
By tradition - and for historical reasons - reflex pathways and interneurones have been named by their dominating sensory input. Later studies have demonstrated that each individual interneurone, as a rule, receives a broad convergence from a large variety of sensory modalities, as well as inputs from one or more descending tracts. It is thus possible that the traditional nomenclature inadvertently has served as a 'straightjacket' for conceptual development in this field. Indeed, there is now much evidence in favour of the view that the many classes of spinal interneurones may be seen as 'functional units' representing different levels of muscle synergies, parts of movements, or even more integrated motor behaviour. Such 'functional units' may be used by (different) descending pathways to mediate the motor commands from the brain and integrate the appropriate (multimodal) sensory feedback into the central command. A given sensory stimulus would then be able to affect the motor output through a number of parallel, or alternative, segmental pathways belonging to different 'functional units'. If this were correct it would indeed be predicted, rather than coming as a surprise, that a given sensory stimulus can result in different outputs - even with a different sign - depending on the preceding selection of active 'functional units', i.e. the type of motor activity initiated by the brain.
Figures
Similar articles
-
Reflex pathways from group II muscle afferents. 3. Secondary spindle afferents and the FRA: a new hypothesis.Exp Brain Res. 1987;65(2):294-306. doi: 10.1007/BF00236301. Exp Brain Res. 1987. PMID: 3556458
-
Common interneurones in reflex pathways from group 1a and 1b afferents of ankle extensors in the cat.J Physiol. 1981 Jan;310:381-402. doi: 10.1113/jphysiol.1981.sp013556. J Physiol. 1981. PMID: 7230041 Free PMC article.
-
[Reflex circuits of the spinal cord in man. Control during movement and their functional role (1)].Rev Neurol (Paris). 1984;140(11):605-14. Rev Neurol (Paris). 1984. PMID: 6239356 French.
-
Spinal functions in sensorimotor control of movements.Neurosurg Rev. 1990;13(3):179-85. doi: 10.1007/BF00313016. Neurosurg Rev. 1990. PMID: 2204840 Review.
-
Interactions between pathways controlling posture and gait at the level of spinal interneurones in the cat.Prog Brain Res. 1993;97:161-71. doi: 10.1016/s0079-6123(08)62274-8. Prog Brain Res. 1993. PMID: 8234742 Review.
Cited by
-
Tapping into rhythm generation circuitry in humans during simulated weightlessness conditions.Front Syst Neurosci. 2015 Feb 18;9:14. doi: 10.3389/fnsys.2015.00014. eCollection 2015. Front Syst Neurosci. 2015. PMID: 25741250 Free PMC article. Review.
-
Convergence in reflex pathways from multiple cutaneous nerves innervating the foot depends upon the number of rhythmically active limbs during locomotion.PLoS One. 2014 Aug 29;9(8):e104910. doi: 10.1371/journal.pone.0104910. eCollection 2014. PLoS One. 2014. PMID: 25170606 Free PMC article.
-
Control of the gain of visual-motor transmission occurs in visual coordinates for smooth pursuit eye movements.J Neurosci. 2013 May 29;33(22):9420-30. doi: 10.1523/JNEUROSCI.4846-12.2013. J Neurosci. 2013. PMID: 23719810 Free PMC article.
-
Spinal interneuronal systems: identification, multifunctional character and reconfigurations in mammals.J Physiol. 2001 May 15;533(Pt 1):31-40. doi: 10.1111/j.1469-7793.2001.0031b.x. J Physiol. 2001. PMID: 11351010 Free PMC article. Review.
-
Whole Body Coordination for Self-Assistance in Locomotion.Front Neurorobot. 2022 Jun 10;16:883641. doi: 10.3389/fnbot.2022.883641. eCollection 2022. Front Neurorobot. 2022. PMID: 35747075 Free PMC article.
References
-
- Zehr EP, Stein RB. What functions do reflexes serve during human locomotion. Progress in Neurobiolology. 1999;58:185–205. - PubMed
-
- Baldissera F, Hultborn H, Illert M. Integration in spinal neuronal systems. In: Brooks VB, editor. Handbook of Physiology The Nervous System Motor Control. II. Bethesda, MD, USA: American Physiological Society; 1981. pp. 509–595. section 1.
-
- Christensen LO, Petersen N, Andersen JB, Sinkjaer T, Nielsen JB. Evidence for transcortical reflex pathways in the lower limb of man. Progress in Neurobiology. 2000;62:251–272. - PubMed
-
- Crone C, Nielsen J. Central control of disynaptic reciprocal inhibition in humans. Acta Physiologica Scandinavica. 1994;152:351–363. - PubMed
-
- Dietz V, Duysens J. Significance of load receptor input during locomotion: a review. Gait and Posture. 2000;11:102–110. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources