

Metaphase arrest with centromere separation in polo mutants of Drosophila
- PMID: 11352929
- PMCID: PMC2192380
- DOI: 10.1083/jcb.153.4.663
Metaphase arrest with centromere separation in polo mutants of Drosophila
Abstract
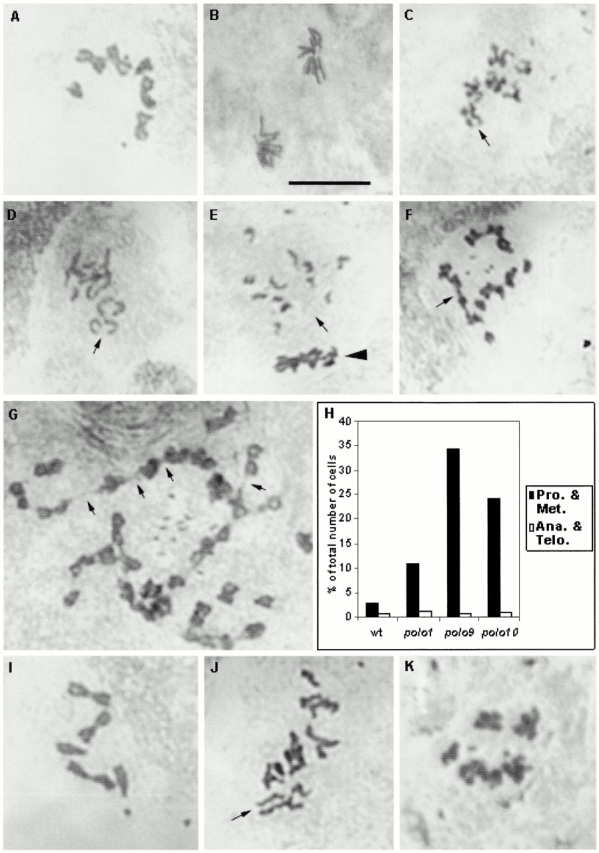
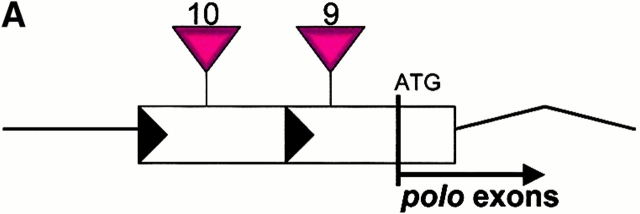
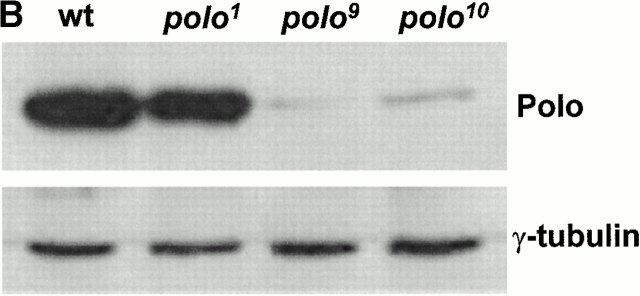
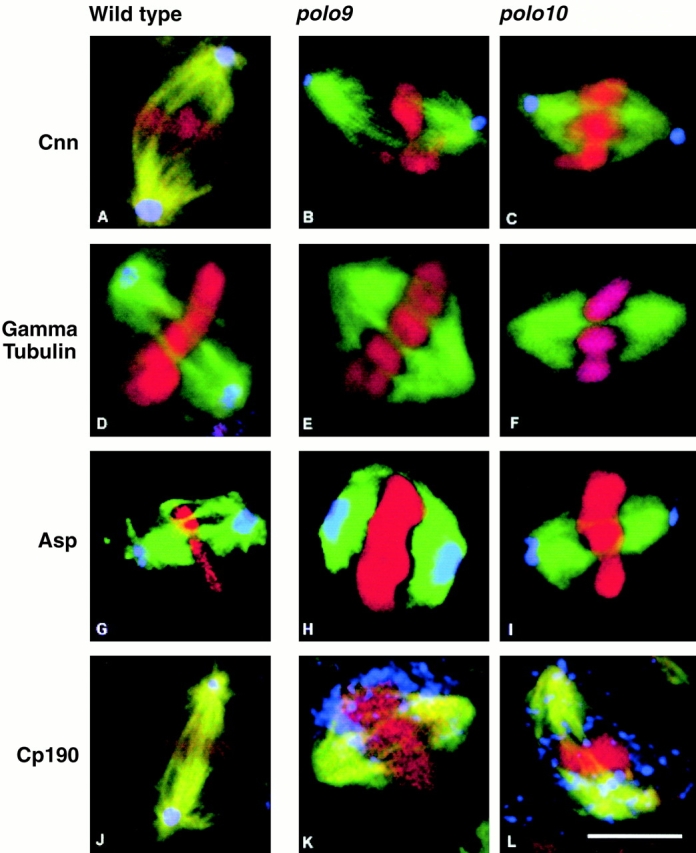
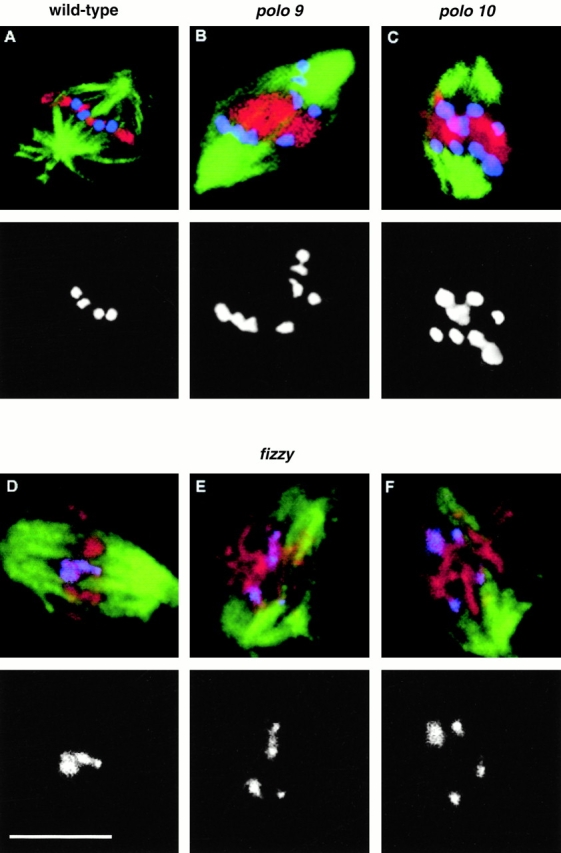
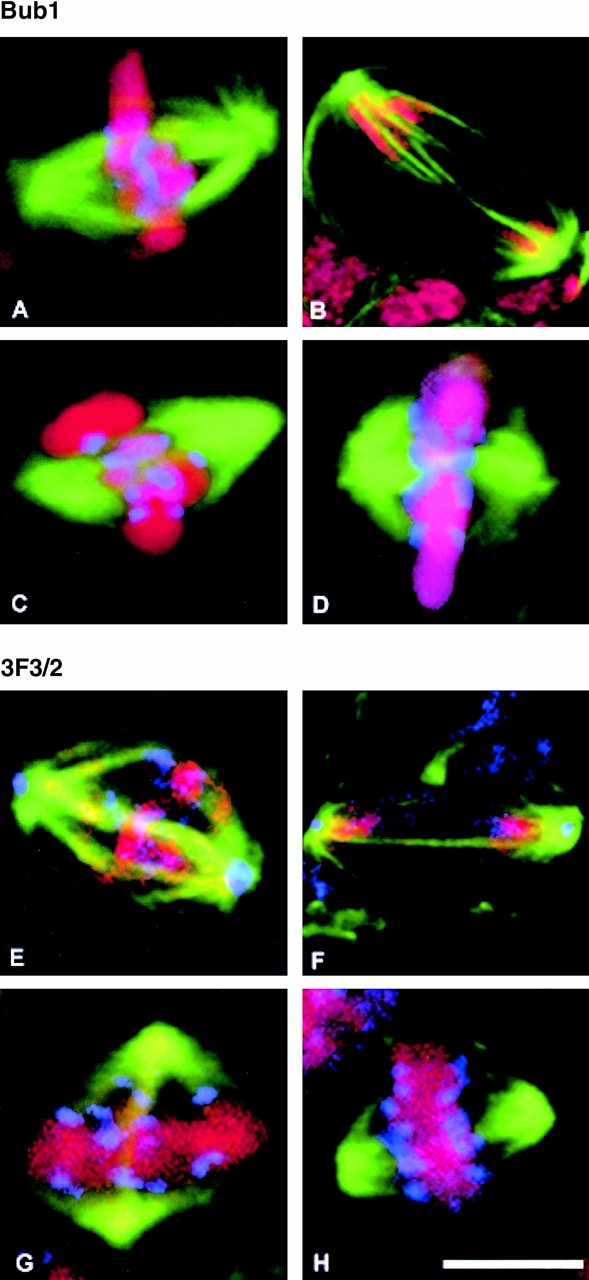
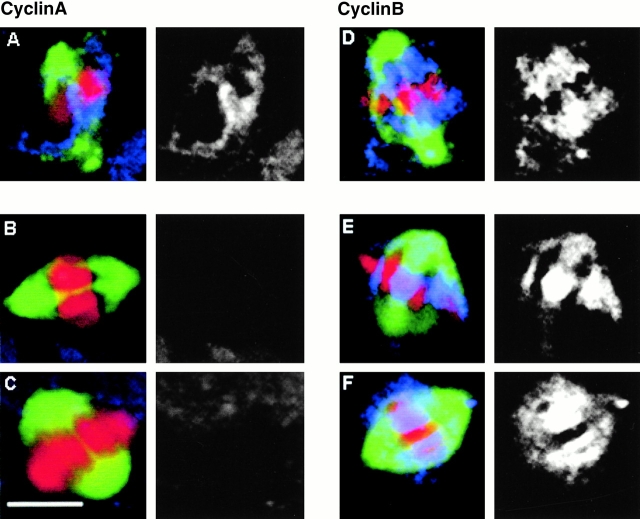
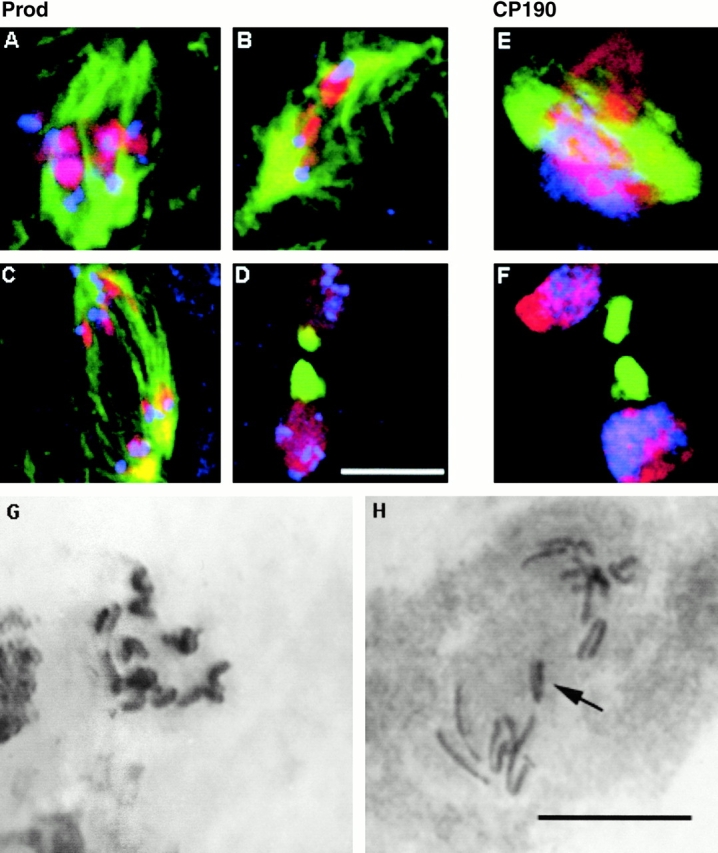
The Drosophila gene polo encodes a conserved protein kinase known to be required to organize spindle poles and for cytokinesis. Here we report two strongly hypomorphic mutations of polo that arrest cells of the larval brain at a point in metaphase when the majority of sister kinetochores have separated by between 20-50% of the total spindle length in intact cells. In contrast, analysis of sister chromatid separation in squashed preparations of cells indicates that some 83% of sisters remain attached. This suggests the separation seen in intact cells requires the tension produced by a functional spindle. The point of arrest corresponds to the spindle integrity checkpoint; Bub1 protein and the 3F3/2 epitope are present on the separated kinetochores and the arrest is suppressed by a bub1 mutation. The mutant mitotic spindles are anastral and have assembled upon centrosomes that are associated with Centrosomin and the abnormal spindle protein (Asp), but neither with gamma-tubulin nor CP190. We discuss roles for Polo kinase in recruiting centrosomal proteins and in regulating progression through the metaphase-anaphase checkpoint.
Figures
References
-
- Abrieu A., Brassac T., Galas S., Fisher D., Labbe J.-C., Doree M. The Polo-like kinase Plx is a component of the MPF amplification loop at the G2/M-phase transition of the cell cycle in Xenopus . J. Cell Sci. 1998;111:1751–1757. - PubMed
-
- Amon A. The spindle checkpoint. Curr. Opin. Genes Dev. 1999;9:69–75. - PubMed
-
- Avides M., Glover D.M. Abnormal spindle protein Asp, and the integrity of mitotic centrosomal microtubule organizing centres. Science. 1999;283:1733–1735. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
