

Phosphorylation-dependent regulation of ryanodine receptors: a novel role for leucine/isoleucine zippers
- PMID: 11352932
- PMCID: PMC2192391
- DOI: 10.1083/jcb.153.4.699
Phosphorylation-dependent regulation of ryanodine receptors: a novel role for leucine/isoleucine zippers
Abstract
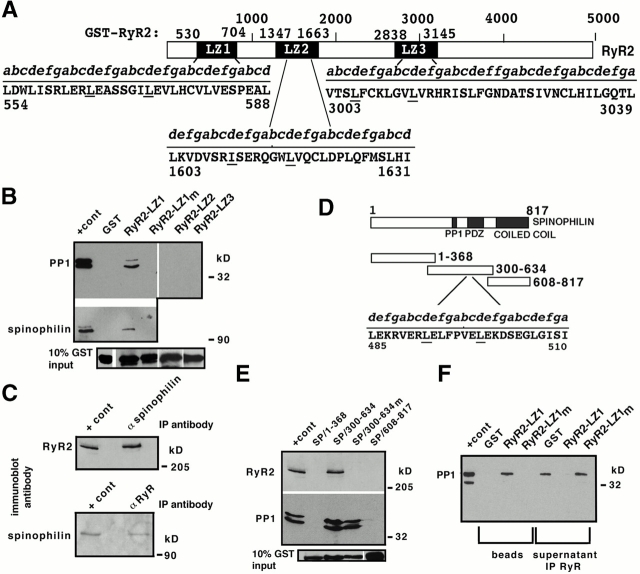
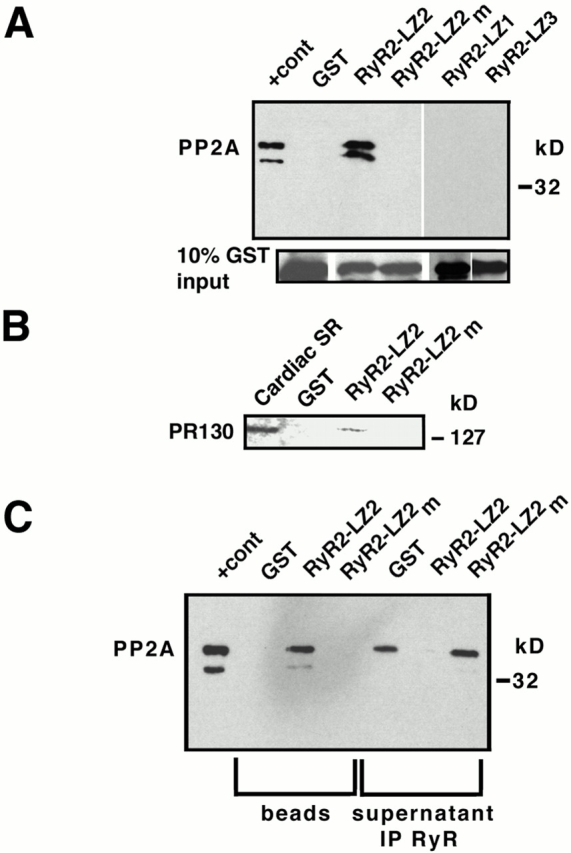
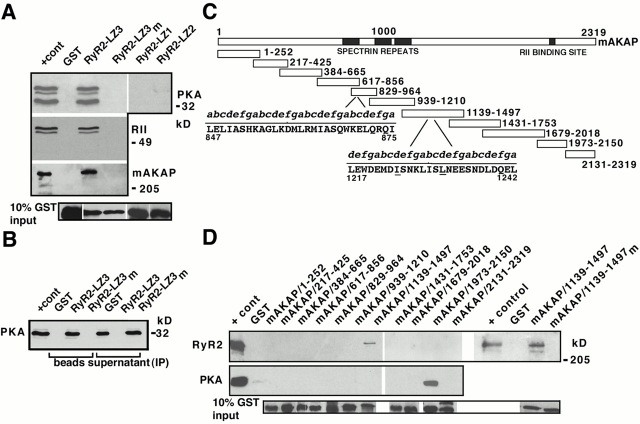
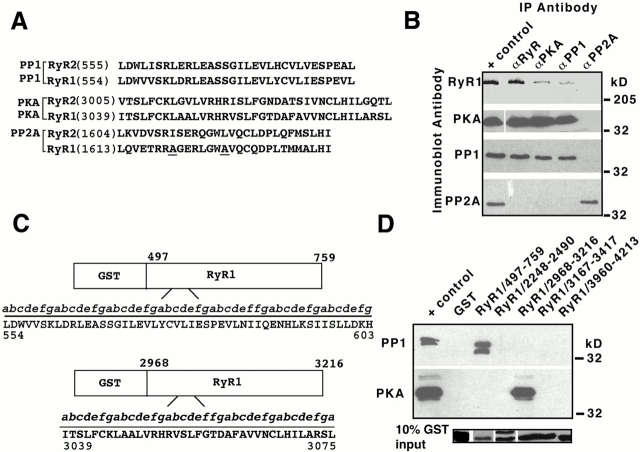
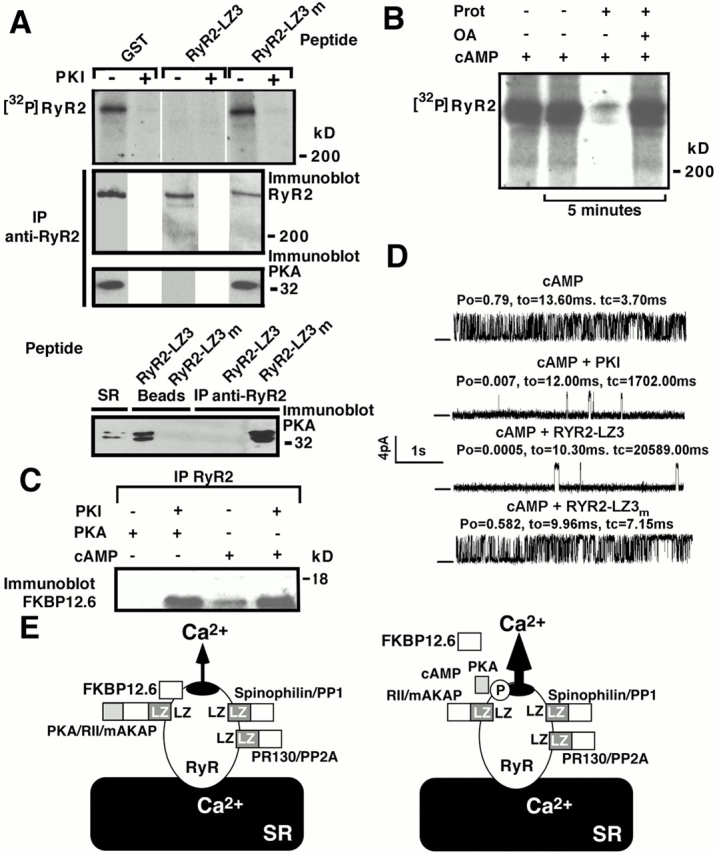
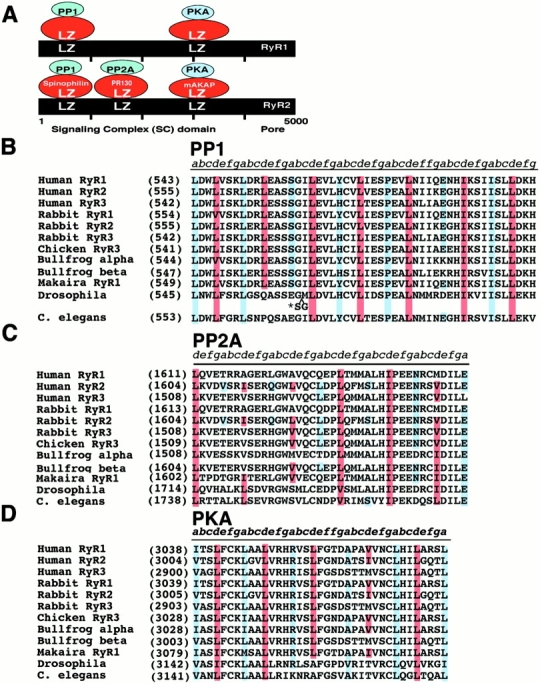
Ryanodine receptors (RyRs), intracellular calcium release channels required for cardiac and skeletal muscle contraction, are macromolecular complexes that include kinases and phosphatases. Phosphorylation/dephosphorylation plays a key role in regulating the function of many ion channels, including RyRs. However, the mechanism by which kinases and phosphatases are targeted to ion channels is not well understood. We have identified a novel mechanism involved in the formation of ion channel macromolecular complexes: kinase and phosphatase targeting proteins binding to ion channels via leucine/isoleucine zipper (LZ) motifs. Activation of kinases and phosphatases bound to RyR2 via LZs regulates phosphorylation of the channel, and disruption of kinase binding via LZ motifs prevents phosphorylation of RyR2. Elucidation of this new role for LZs in ion channel macromolecular complexes now permits: (a) rapid mapping of kinase and phosphatase targeting protein binding sites on ion channels; (b) predicting which kinases and phosphatases are likely to regulate a given ion channel; (c) rapid identification of novel kinase and phosphatase targeting proteins; and (d) tools for dissecting the role of kinases and phosphatases as modulators of ion channel function.
Figures
References
-
- Alber T., Bell J.A., Sun D.P., Nicholson H., Wozniak J.A., Cook S., Matthews B.W. Replacements of Pro86 in phage T4 lysozyme extend an alpha-helix but do not alter protein stability. Science. 1988;239:631–635. - PubMed
-
- Angelo R., Rubin C.S. Molecular characterization of an anchor protein (AKAPCE) that binds the RI subunit (RCE) of type I protein kinase A from Caenorhabditis elegans . J. Biol. Chem. 1998;273:14633–14643. - PubMed
-
- Barlow D.J., Thornton J.M. Helix geometry in proteins. J. Mol. Biol. 1988;201:601–619. - PubMed
-
- Brillantes A.B., Ondrias K., Scott A., Kobrinsky E., Ondriasova E., Moschella M.C., Jayaraman T., Landers M., Ehrlich B.E., Marks A.R. Stabilization of calcium release channel (ryanodine receptor) function by FK506-binding protein. Cell. 1994;77:513–523. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
