

doi: 10.1093/nar/29.10.2041.
Antisense oligonucleotides selected by hybridisation to scanning arrays are effective reagents in vivo
Affiliations
- PMID: 11353073
- PMCID: PMC55457
- DOI: 10.1093/nar/29.10.2041
Item in Clipboard
Antisense oligonucleotides selected by hybridisation to scanning arrays are effective reagents in vivo
Nucleic Acids Res.
.
Abstract
Transcripts representing mRNAs of three Xenopus cyclins, B1, B4 and B5, were hybridised to arrays of oligonucleotides scanning the first 120 nt of the coding region to assess the ability of the immobilised oligonucleotides to form heteroduplexes with their targets. Oligonucleotides that produced high heteroduplex yield and others that showed little annealing were assayed for their effect on translation of endogenous cyclin mRNAs in Xenopus egg extracts and their ability to promote cleavage of cyclin mRNAs in oocytes by RNase H. Excellent correlation was found between antisense potency and affinity of oligonucleotides for the cyclin transcripts as measured by the array, despite the complexity of the cellular environment.
Figures
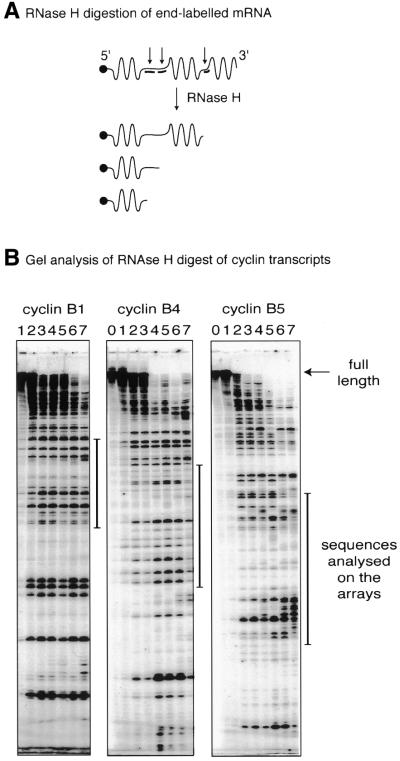
Identification of accessible
regions on the cyclin transcripts with RNase H/dN12 (A) Schematic representation of RNase H digestion
of end-labelled mRNA. (B) Autoradiographs showing
gel analysis of the cleaved products of 5′-end-labelled
mRNA with RNase H at various dN12 concentrations. The
transcripts were titrated against various concentrations of the
oligomer library, ranging from 10 to 500 pmol/reaction
(lane 0, native transcript; lane 1, plus RNase H; lane 2, 10 pmol;
lane 3, 20 pmol; lane 4, 50 pmol; lane 5, 100 pmol; lane 6, 250
pmol; lane 7, 500 pmol). Each reaction contained ∼3 × 109 transcript molecules.
The dN12 library contains 1.67 × 107 different
12mers. There are ∼1.8 × 107 molecules
of each oligonucleotide in 500 pmol, which resulted in almost complete
cutting of the full-length transcripts. How can this number of oligonucleotides
mediate cleavage of 3 × 109 target
molecules? Since regions of complementarity between RNA and an oligonucleotide
of only 4 nt can recruit RNase H activity (25), the oligonucleotides
in the library that were only partially complementary to the accessible sites
would also be expected to participate in RNase H-mediated cutting
of the transcript. In addition, each oligonucleotide can participate
more than once in successive reactions.
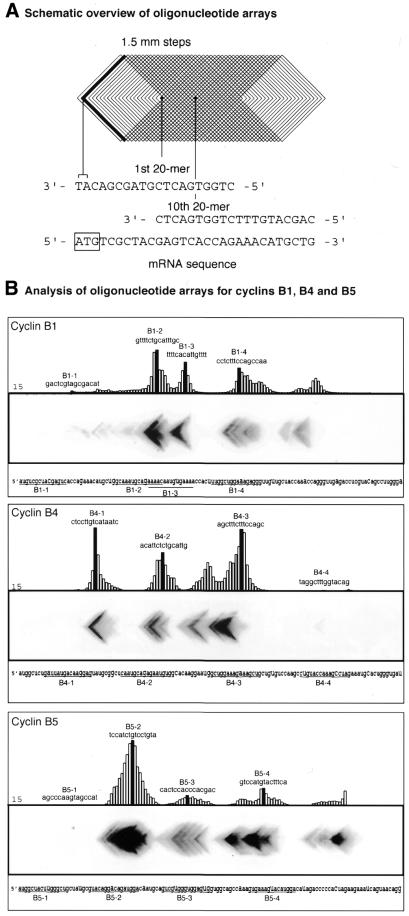
The effects on binding capacity
of sequences and lengths of antisense oligonculeotides determined
on scanning arrays. (A) Schematic representation
of a scanning array. The sequences given in the cartoon correspond
to cyclin B5. (B) Each of the three panels shows
an image obtained after hybridisation of the radiolabelled transcripts
to the corresponding scanning array. The histograms, obtained with
the program xvseq, represent binding of the 15mer oligonucleotides to
the transcript on the array. Histograms corresponding to only half
of the arrays are shown. The sequences of the antisense oligonucleotides
used in various assays are shown on the histograms and underlined
in the 120 nt sequence of the cyclin mRNAs.
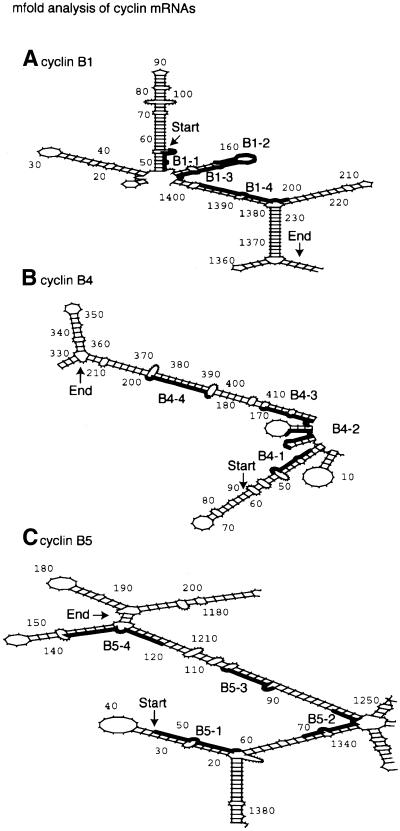
Folded structures of the
three cyclin mRNAs obtained using mfold. Only the relevant sections
of the folds are shown. The sequences analysed on arrays are marked
(start and end are indicated). The binding sites of the oligonucleotides
are marked with bold lines on each structure. The grey line in the
B1 mRNA fold represents overlap between oligonucleotides B1-2 and
B1-3.
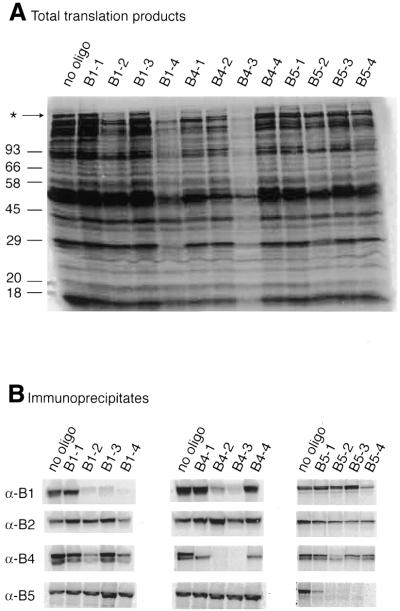
Effects of oligonucleotides
on cell-free protein synthesis in frog egg extract. (A)
The pattern of radioactive proteins displayed on SDS–PAGE
after addition of 1 µM of the indicated
oligonucleotides. The asterisk indicates an unidentified high molecular
weight translation product whose synthesis is particularly sensitive
to non-specific inhibition by the added DNA. (B)
The radiolabelled immunoprecipitates obtained with the indicated
antisera in response to the various oligonucleotides.
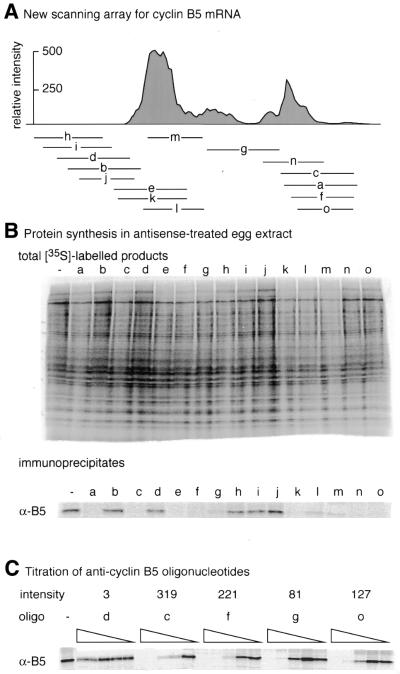
(A) Designation,
location and hybridisation intensities of the oligonucleotides tested
for their effects on translation in (B), which
shows both the total pattern of protein synthesis and the specific
synthesis of B5 using immunoprecipitation and autoradiography. (C) Titration of selected oligonucleotides from
2 to 0.125 µM in five 2-fold dilution
steps as indicated by the wedges. Note that oligonucleotides c,
f and o cover the same region of the mRNA, but differ in length
and hybridisation intensity as indicated. Oligonucleotide d comes
from a poorly scoring region of the array.
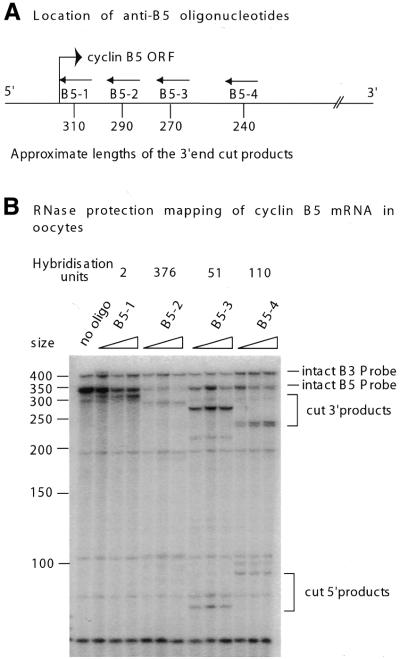
Cleavage of cyclin B5 mRNA
by antisense 15mers injected into intact Xenopus oocytes.
(A) The location of four antisense oligonucleotides
on the ∼350 nt probe and approximate
sizes of the 3′-end cut products (drawings
only roughly to scale). (B) The RNase protection
map for cyclin B5 mRNA in response to antisense-mediated cleavage
produced by 25, 50 and 100 ng each oligonucleotide as indicated
by the wedges. Cyclin B3 was used as a negative control and the products
of cleavage are indicated on the right side of the figure. The additional
fainter bands seen in the B5-3 and B5-4 lanes possibly represent
further degradation products of the B5 mRNA after cleavage with
antisense oligonucleotides, since the oligonucleotides do not have
sufficient complementarity in the neighbouring regions (data not
shown) for secondary binding to produce faint bands of these sizes.
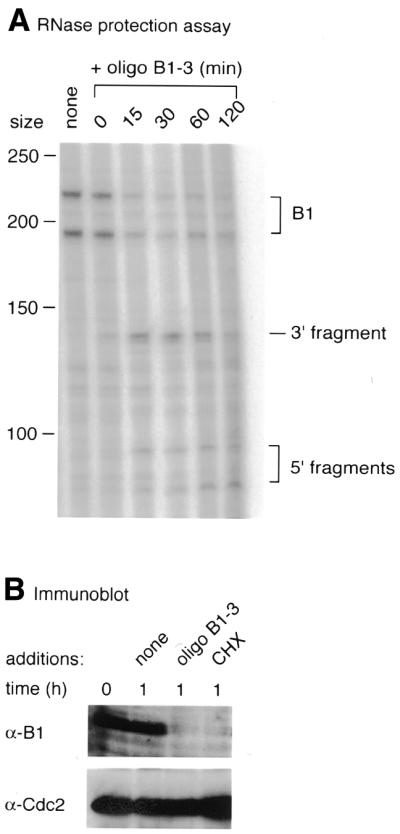
Time course of cleavage of
cyclin B1 mRNA and effects on the levels of cyclin B1 protein in
response to oligonucleotide B1-3. (A) Cleavage
of cyclin B1 mRNA as measured by RNase protection mapping at the
indicated times after injection of the DNA into progesterone-treated Xenopus oocytes. (B) Immunoblots
of cyclin B1 and Cdc2 (loading control) in progesterone-treated
oocytes at the time of GVBD (0) or 1 h later after injection of
water or oligonucleotide solution as indicated or addition of 100 µg/ml cycloheximide (CHX) to
the medium.
References
-
- Branch A.D. (1998) Antisense drug discovery: can cell-free screens speed the process? Antisense Nucleic Acid Drug Dev., 8, 249–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
