

Neural induction in the absence of mesoderm: beta-catenin-dependent expression of secreted BMP antagonists at the blastula stage in Xenopus
- PMID: 11356027
- PMCID: PMC3039525
- DOI: 10.1006/dbio.2001.0258
Neural induction in the absence of mesoderm: beta-catenin-dependent expression of secreted BMP antagonists at the blastula stage in Xenopus
Abstract
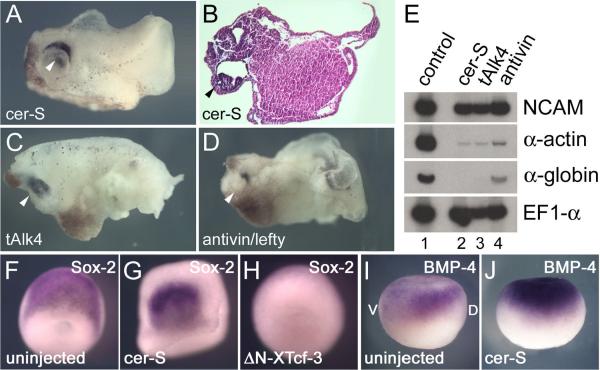
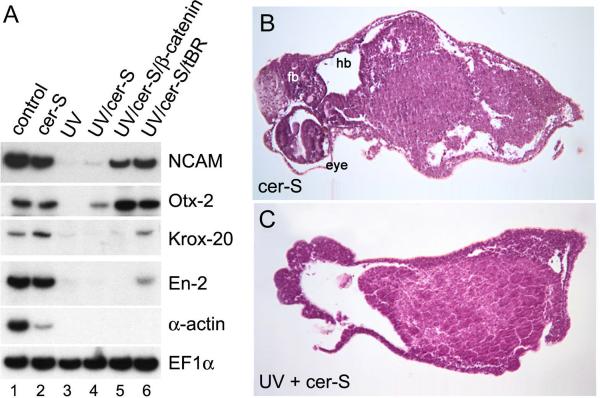
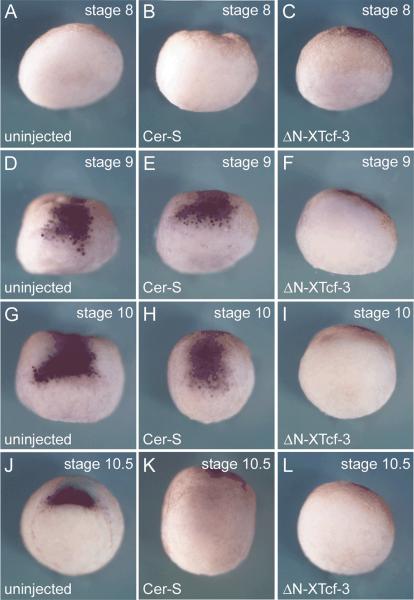
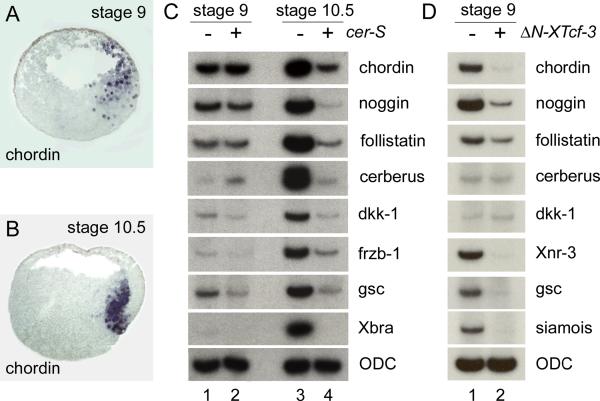
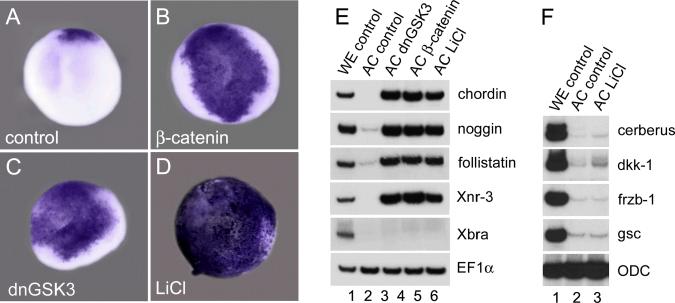
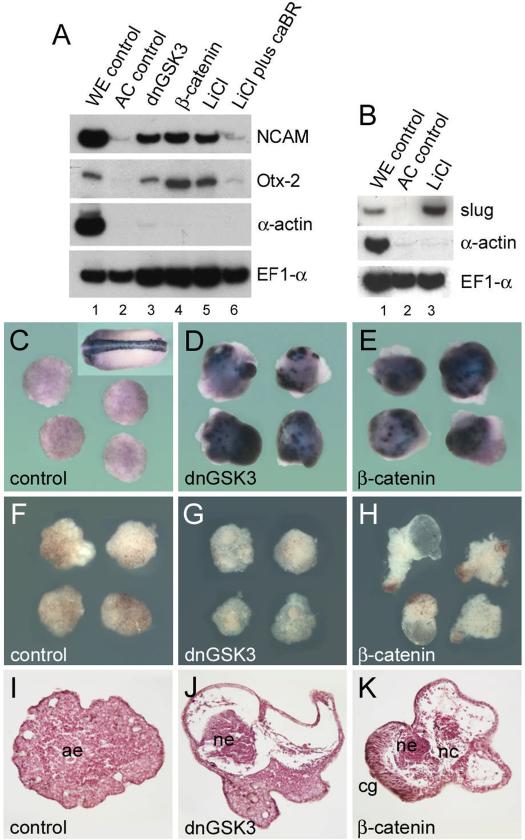
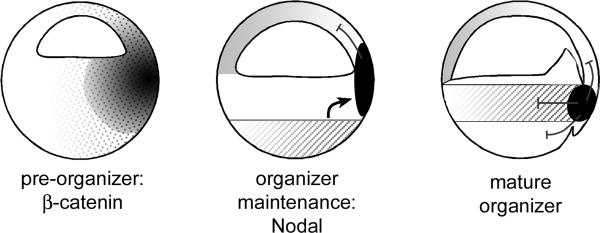
A growing body of work indicates that neural induction may be initiated prior to the establishment of the gastrula mesodermal organizer. Here, we examine neural induction in Xenopus embryos in which mesoderm induction has been blocked by Cerberus-short, a reagent that specifically inhibits Nodal-related (Xnr) signals. We find that extensive neural structures with cyclopic eyes and brain tissue are formed despite the absence of mesoderm. This neural induction correlates with the expression of chordin and other BMP inhibitors-such as noggin, follistatin, and Xnr3-at the blastula stage, and requires beta-Catenin signaling. Activation of the beta-Catenin pathway by mRNA microinjections or by treatment with LiCl leads to differentiation of neurons, as well as neural crest, in ectodermal explants. Xnr signals are required for the maintenance, but not for the initiation, of BMP antagonist expression. Recent work has demonstrated a role for beta-Catenin signaling in neural induction mediated by the transcriptional down-regulation of BMP-4 expression. The present results suggest an additional function for beta-Catenin, the early activation of expression of secreted BMP antagonists, such as Chordin, in a preorganizer region in the dorsal side of the Xenopus blastula.
Copyright 2001 Academic Press.
Figures
Similar articles
-
Neural induction in Xenopus: requirement for ectodermal and endomesodermal signals via Chordin, Noggin, beta-Catenin, and Cerberus.PLoS Biol. 2004 May;2(5):E92. doi: 10.1371/journal.pbio.0020092. Epub 2004 May 11. PLoS Biol. 2004. PMID: 15138495 Free PMC article.
-
Dorsal-ventral patterning and neural induction in Xenopus embryos.Annu Rev Cell Dev Biol. 2004;20:285-308. doi: 10.1146/annurev.cellbio.20.011403.154124. Annu Rev Cell Dev Biol. 2004. PMID: 15473842 Free PMC article. Review.
-
Depletion of three BMP antagonists from Spemann's organizer leads to a catastrophic loss of dorsal structures.Dev Cell. 2005 Mar;8(3):401-11. doi: 10.1016/j.devcel.2005.01.013. Dev Cell. 2005. PMID: 15737935
-
Anterior endomesoderm specification in Xenopus by Wnt/beta-catenin and TGF-beta signalling pathways.Dev Biol. 1999 May 15;209(2):282-97. doi: 10.1006/dbio.1999.9257. Dev Biol. 1999. PMID: 10328921
-
[Mesoderm induction and neural induction].Tanpakushitsu Kakusan Koso. 2005 May;50(6 Suppl):583-7. Tanpakushitsu Kakusan Koso. 2005. PMID: 15926484 Review. Japanese. No abstract available.
Cited by
-
Dynamic in vivo binding of transcription factors to cis-regulatory modules of cer and gsc in the stepwise formation of the Spemann-Mangold organizer.Development. 2012 May;139(9):1651-61. doi: 10.1242/dev.068395. Development. 2012. PMID: 22492356 Free PMC article.
-
POU-V factors antagonize maternal VegT activity and beta-Catenin signaling in Xenopus embryos.EMBO J. 2007 Jun 20;26(12):2942-54. doi: 10.1038/sj.emboj.7601736. Epub 2007 May 31. EMBO J. 2007. PMID: 17541407 Free PMC article.
-
xBtg-x regulates Wnt/beta-Catenin signaling during early Xenopus development.Dev Biol. 2005 Jul 1;283(1):17-28. doi: 10.1016/j.ydbio.2005.03.033. Dev Biol. 2005. PMID: 15975429 Free PMC article.
-
Rap2 is required for Wnt/beta-catenin signaling pathway in Xenopus early development.EMBO J. 2005 Mar 9;24(5):985-96. doi: 10.1038/sj.emboj.7600571. Epub 2005 Feb 10. EMBO J. 2005. PMID: 15706349 Free PMC article.
-
Neural plate patterning by secreted signals.Neuron. 2002 Feb 14;33(4):489-91. doi: 10.1016/s0896-6273(02)00596-2. Neuron. 2002. PMID: 11856521 Free PMC article. Review.
References
-
- Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, May SR, McMahon JA, McMahon AP, Harland RM, Rossant J, De Robertis EM. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature. 2000;403:658–661. - PubMed
-
- Bauer DV, Huang S, Moody SA. The cleavage stage origin of Spemann's Organizer: analysis of the movements of blastomere clones before and during gastrulation in Xenopus. Development. 1994;120:1179–1189. - PubMed
-
- Belo JA, Bouwmeester T, Leyns L, Kertesz N, Gallo M, Follettie M, De Robertis EM. Cerberus-like is a secreted factor with neutralizing activity expressed in the anterior primitive endoderm of the mouse gastrula. Mech. Dev. 1997;68:45–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
