

Metabotropic glutamate receptor subtypes 1 and 5 are activators of extracellular signal-regulated kinase signaling required for inflammatory pain in mice
- PMID: 11356865
- PMCID: PMC6762705
- DOI: 10.1523/JNEUROSCI.21-11-03771.2001
Metabotropic glutamate receptor subtypes 1 and 5 are activators of extracellular signal-regulated kinase signaling required for inflammatory pain in mice
Abstract
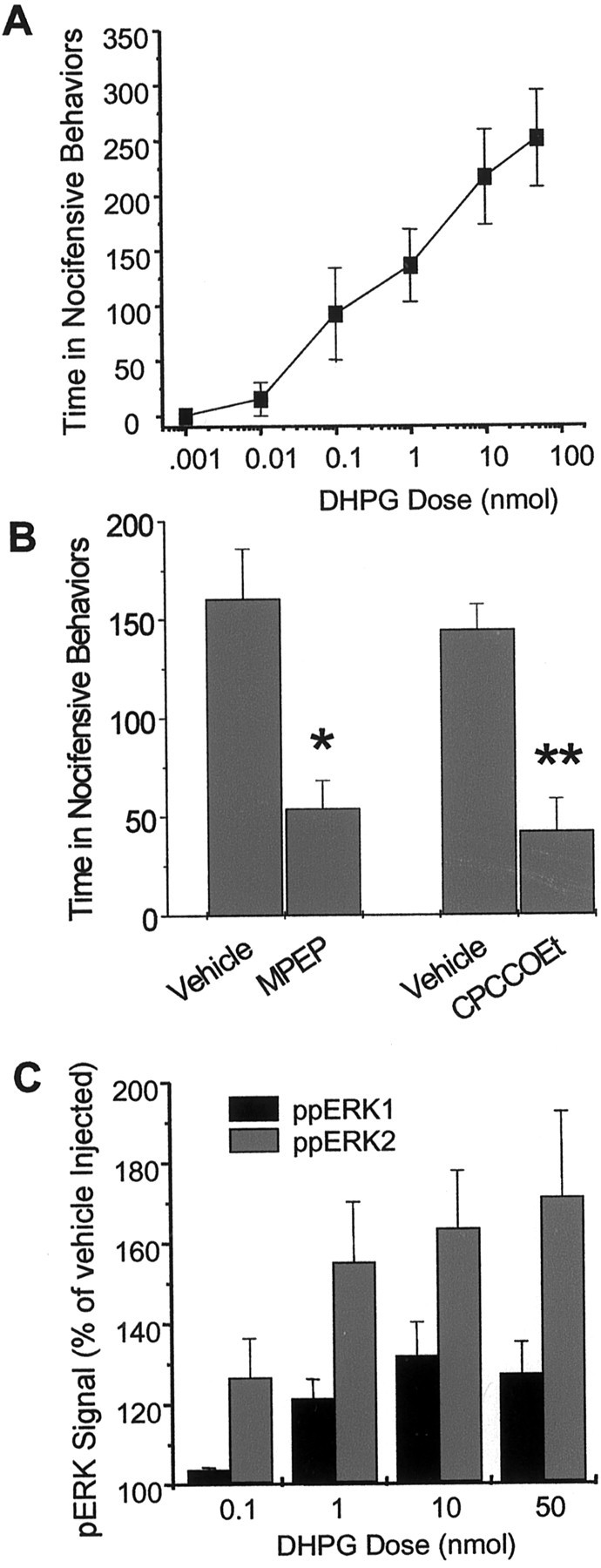
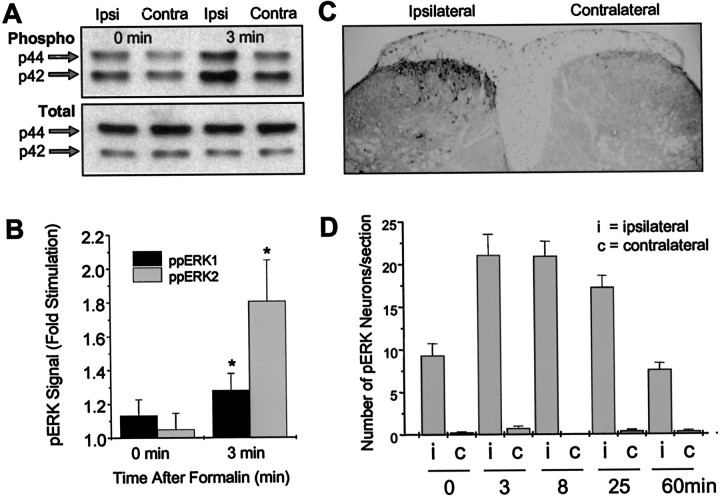
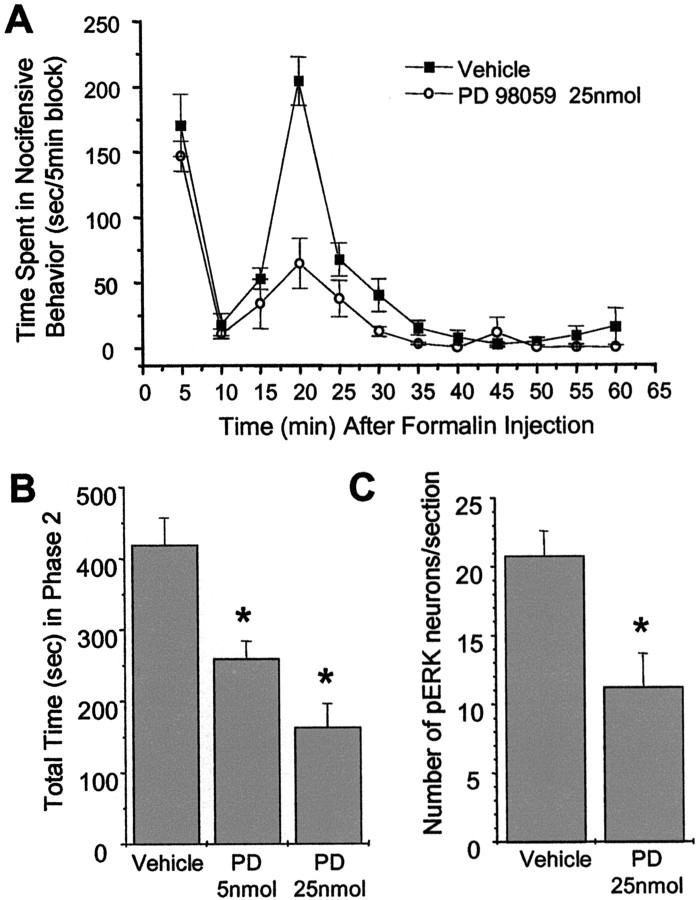
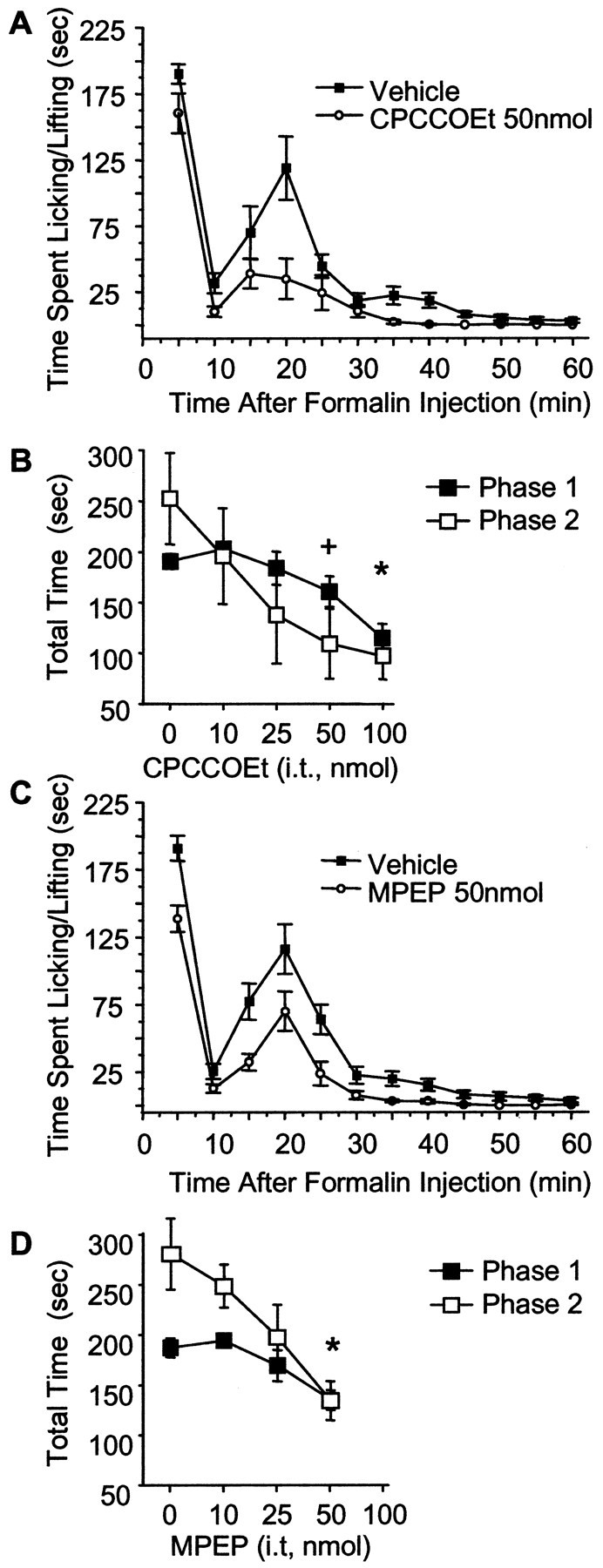
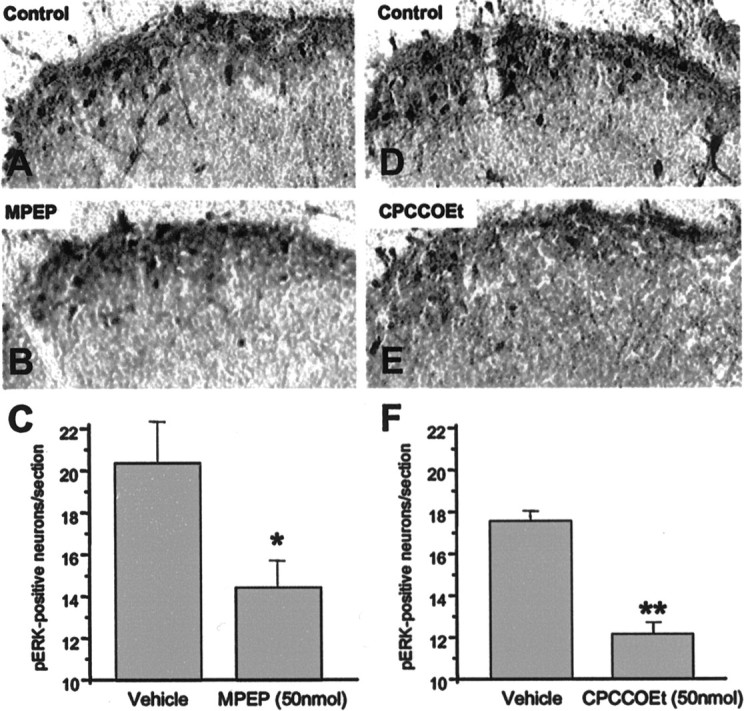
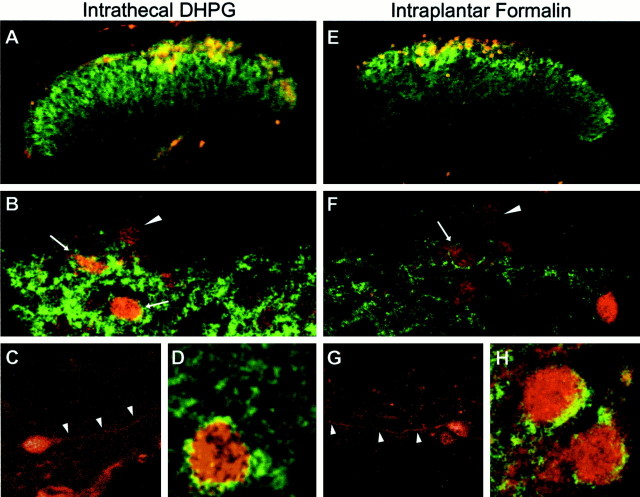
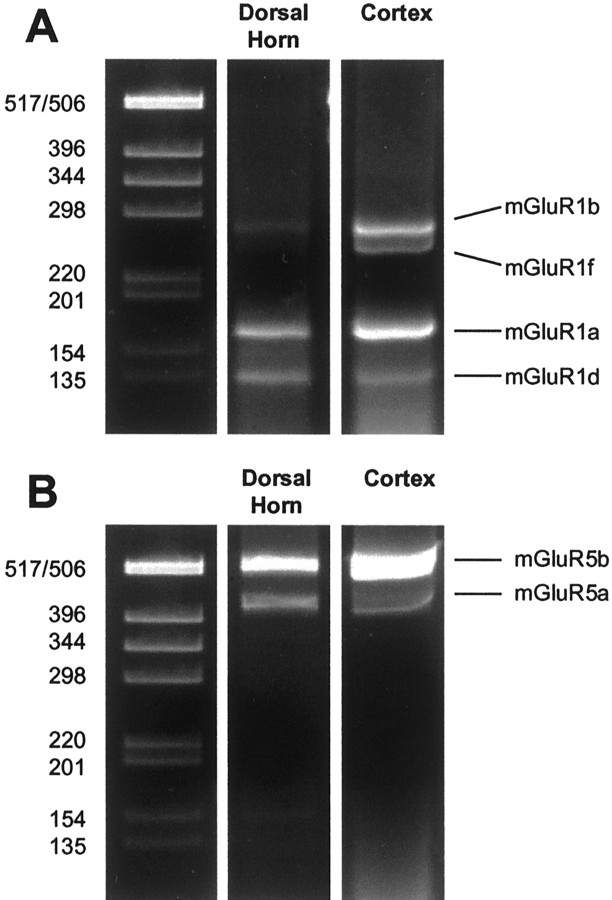
Metabotropic glutamate receptors are expressed abundantly in the spinal cord and have been shown to play important roles in the modulation of nociceptive transmission and plasticity. Most previous studies have focused on the group I metabotropic glutamate receptors (mGluR1 and mGluR5) and activation of phospholipase C signaling by these receptors in modulating nociception. Recently, it was shown that the extracellular signal-regulated kinases (ERKs)/mitogen-activated protein kinases are activated in spinal cord dorsal horn neurons in response to stimulation of nociceptors and that ERK signaling is involved in nociceptive plasticity. In the present studies, we sought to test the hypothesis that group I mGluRs modulate nociceptive transmission or plasticity via modulation of ERK signaling in dorsal horn neurons. We show that activation of mGluR1 and mGluR5 leads to activation of ERK1 and ERK2 in the spinal cord. Furthermore, we find that inflammation-evoked ERK activation, which is required for nociceptive plasticity, is downstream of mGluR1 and mGluR5. Finally, we show colocalization of group I mGluRs with activated ERK in dorsal horn neurons. These results show that mGluR1 and mGluR5 are activated in dorsal horn neurons in response to peripheral inflammation and that activation of these group I mGluRs leads to activation of ERK1 and ERK2, resulting in enhanced pain sensitivity.
Figures
References
-
- Alvarez FJ, Villalba RM, Carr PA, Grandes P, Somohano PM. Differential distribution of metabotropic glutamate receptors 1a, 1b, and 5 in the rat spinal cord. J Comp Neurol. 2000;422:464–487. - PubMed
-
- Bordi F, Ugolini A. Involvement of mGluR(5) on acute nociceptive transmission. Brain Res. 2000;871:223–233. - PubMed
-
- Boxall SJ, Thompson SW, Dray A, Dickenson AH, Urban L. Metabotropic glutamate receptor activation contributes to nociceptive reflex activity in the rat spinal cord in vitro. Neuroscience. 1996;74:13–20. - PubMed
-
- Budai D, Larson AA. The involvement of metabotropic glutamate receptors in sensory transmission in dorsal horn of the rat spinal cord. Neuroscience. 1998;83:571–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous