

Impaired heme binding and aggregation of mutant cystathionine beta-synthase subunits in homocystinuria
- PMID: 11359213
- PMCID: PMC1226138
- DOI: 10.1086/320597
Impaired heme binding and aggregation of mutant cystathionine beta-synthase subunits in homocystinuria
Abstract
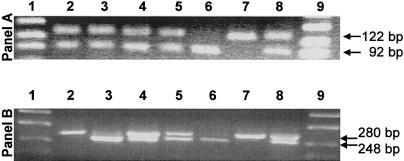
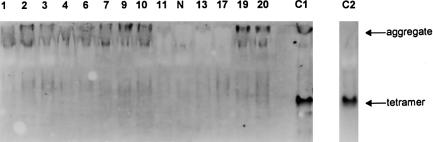
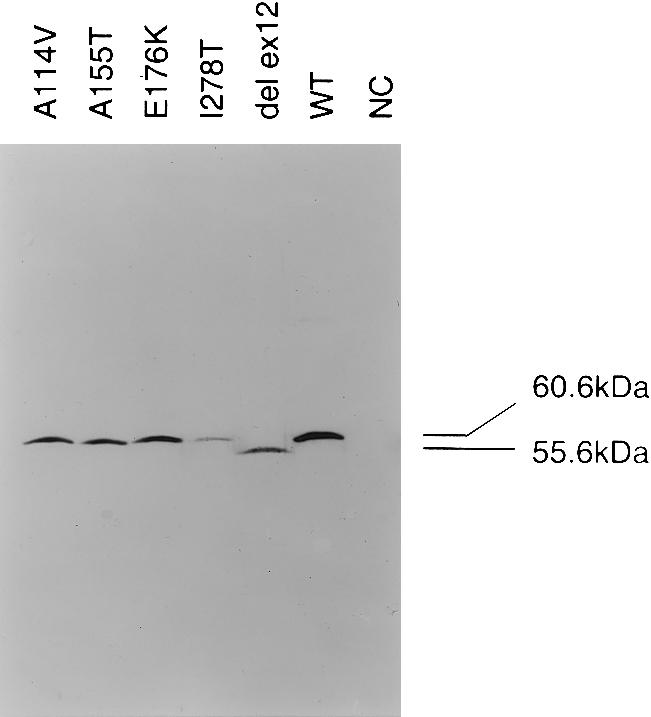
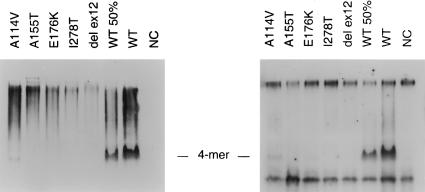
During the past 20 years, cystathionine beta-synthase (CBS) deficiency has been detected in the former Czechoslovakia with a calculated frequency of 1:349,000. The clinical manifestation was typical of homocystinuria, and about half of the 21 patients were not responsive to pyridoxine. Twelve distinct mutations were detected in 30 independent homocystinuric alleles. One half of the alleles carried either the c.833 T-->C or the IVS11-2A-->C mutation; the remaining alleles contained private mutations. The abundance of five mutant mRNAs with premature stop codons was analyzed by PCR-RFLP. Two mRNAs, c.828_931ins104 (IVS7+1G-->A) and c.1226 G-->A, were severely reduced in the cytoplasm as a result of nonsense-mediated decay. In contrast, the other three mRNAs-c.19_20insC, c.28_29delG, and c.210_235del26 (IVS1-1G-->C)-were stable. Native western blot analysis of 14 mutant fibroblast lines showed a paucity of CBS antigen, which was detectable only in aggregates. Five mutations-A114V (c.341C-->T), A155T (c.463G-->A), E176K (c.526G-->A), I278T (c.833T-->C), and W409_G453del (IVS11-2A-->C)-were expressed in Escherichia coli. All five mutant proteins formed substantially more aggregates than did the wild-type CBS, and no aggregates contained heme. These data suggest that abnormal folding, impaired heme binding, and aggregation of mutant CBS polypeptides may be common pathogenic mechanisms in CBS deficiency.
Figures
Similar articles
-
Four novel mutations in the cystathionine beta-synthase gene: effect of a second linked mutation on the severity of the homocystinuric phenotype.Hum Mutat. 1999;13(6):453-7. doi: 10.1002/(SICI)1098-1004(1999)13:6<453::AID-HUMU4>3.0.CO;2-K. Hum Mutat. 1999. PMID: 10408774
-
Reduced response of Cystathionine Beta-Synthase (CBS) to S-Adenosylmethionine (SAM): Identification and functional analysis of CBS gene mutations in Homocystinuria patients.J Inherit Metab Dis. 2014 Mar;37(2):245-54. doi: 10.1007/s10545-013-9647-6. Epub 2013 Aug 23. J Inherit Metab Dis. 2014. PMID: 23974653
-
Identical genotypes in siblings with different homocystinuric phenotypes: identification of three mutations in cystathionine beta-synthase using an improved bacterial expression system.Hum Mol Genet. 1994 Jul;3(7):1103-8. doi: 10.1093/hmg/3.7.1103. Hum Mol Genet. 1994. PMID: 7981678
-
Cystathionine beta-synthase mutations in homocystinuria.Hum Mutat. 1999;13(5):362-75. doi: 10.1002/(SICI)1098-1004(1999)13:5<362::AID-HUMU4>3.0.CO;2-K. Hum Mutat. 1999. PMID: 10338090 Review.
-
A revisit to the natural history of homocystinuria due to cystathionine beta-synthase deficiency.Mol Genet Metab. 2010 Jan;99(1):1-3. doi: 10.1016/j.ymgme.2009.09.009. Mol Genet Metab. 2010. PMID: 19819175 Free PMC article. Review.
Cited by
-
Surrogate genetics and metabolic profiling for characterization of human disease alleles.Genetics. 2012 Apr;190(4):1309-23. doi: 10.1534/genetics.111.137471. Epub 2012 Jan 20. Genetics. 2012. PMID: 22267502 Free PMC article.
-
Cystathionine beta-synthase mutants exhibit changes in protein unfolding: conformational analysis of misfolded variants in crude cell extracts.J Inherit Metab Dis. 2012 May;35(3):469-77. doi: 10.1007/s10545-011-9407-4. Epub 2011 Nov 9. J Inherit Metab Dis. 2012. PMID: 22069143 Free PMC article.
-
Domain organization, catalysis and regulation of eukaryotic cystathionine beta-synthases.PLoS One. 2014 Aug 14;9(8):e105290. doi: 10.1371/journal.pone.0105290. eCollection 2014. PLoS One. 2014. PMID: 25122507 Free PMC article.
-
Purification, crystallization and preliminary crystallographic analysis of human cystathionine β-synthase.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2012 Nov 1;68(Pt 11):1318-22. doi: 10.1107/S1744309112037219. Epub 2012 Oct 30. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2012. PMID: 23143240 Free PMC article.
-
Cystathionine β-synthase is involved in cysteine biosynthesis and H2S generation in Toxoplasma gondii.Sci Rep. 2020 Sep 4;10(1):14657. doi: 10.1038/s41598-020-71469-x. Sci Rep. 2020. PMID: 32887901 Free PMC article.
References
Electronic-Database Information
-
- Authors' Web site, http://www.uchsc.edu/sm/cbs (for an updated list of mutations)
-
- Genbank, http://www.ncbi.nlm.nih.gov/Genbank (for human CBS cDNA [accession number L19501] and genomic DNA [accession number AF042836])
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for CBS deficiency [MIM 236200])
-
- Primers and Conditions, http://www.lf1.cuni.cz/~mjano/protocols.html (for list of PCR primers and conditions for amplification of all 23 CBS exons from genomic DNA)
References
-
- Bross P, Corydon TJ, Andresen BS, Jorgensen MM, Bolund L, Gregersen N (1999) Protein misfolding and degradation in genetic diseases. Hum Mutat 14:186–198 - PubMed
-
- Cooper DN, Krawczak M, Antonarakis SE (1995) The nature and mechanisms of human gene mutation. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The metabolic and molecular bases of inherited disease. 7th ed. McGraw-Hill, New York, pp 259–292
-
- Dawson PA, Cox AJ, Emmerson BT, Dudman NP, Kraus JP, Gordon RB (1997) Characterisation of five missense mutations in the cystathionine beta-synthase gene from three patients with B6-nonresponsive homocystinuria. Eur J Hum Genet 5:15–21 - PubMed
-
- de Franchis R, Kozich V, McInnes RR, Kraus JP (1994) Identical genotypes in siblings with different homocystinuric phenotypes: identification of three mutations in cystathionine beta-synthase using an improved bacterial expression system. Hum Mol Genet 3:1103–1108 - PubMed
-
- Farabaugh PJ (2000) Translational frameshifting: implications for the mechanism of translational frame maintenance. Prog Nucleic Acid Res Mol Biol 64:131–170 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
