

Rag-1 mutations associated with B-cell-negative scid dissociate the nicking and transesterification steps of V(D)J recombination
- PMID: 11359901
- PMCID: PMC87056
- DOI: 10.1128/MCB.21.12.3935-3946.2001
Rag-1 mutations associated with B-cell-negative scid dissociate the nicking and transesterification steps of V(D)J recombination
Abstract
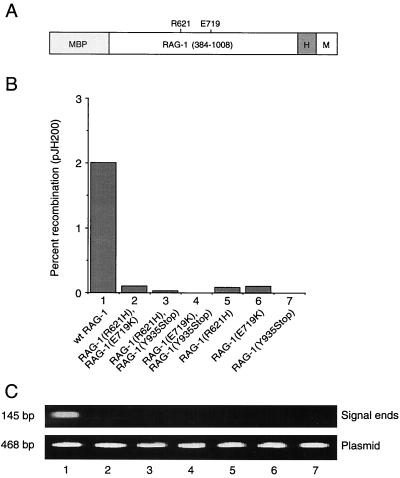
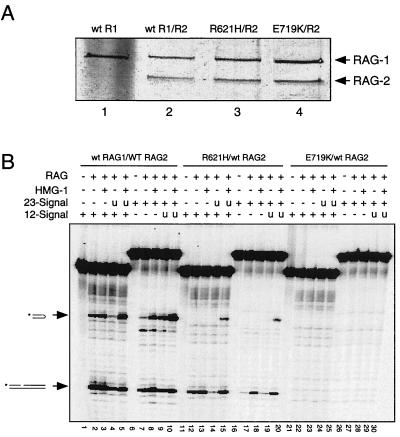
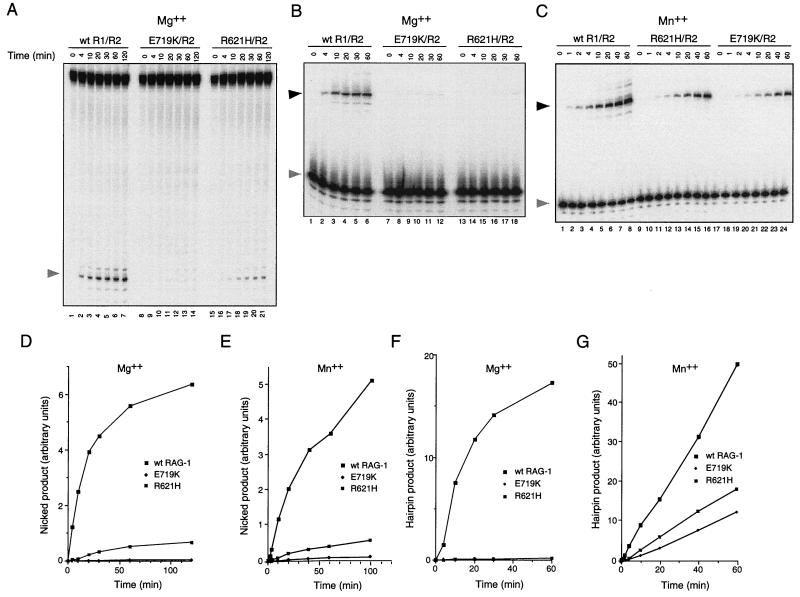
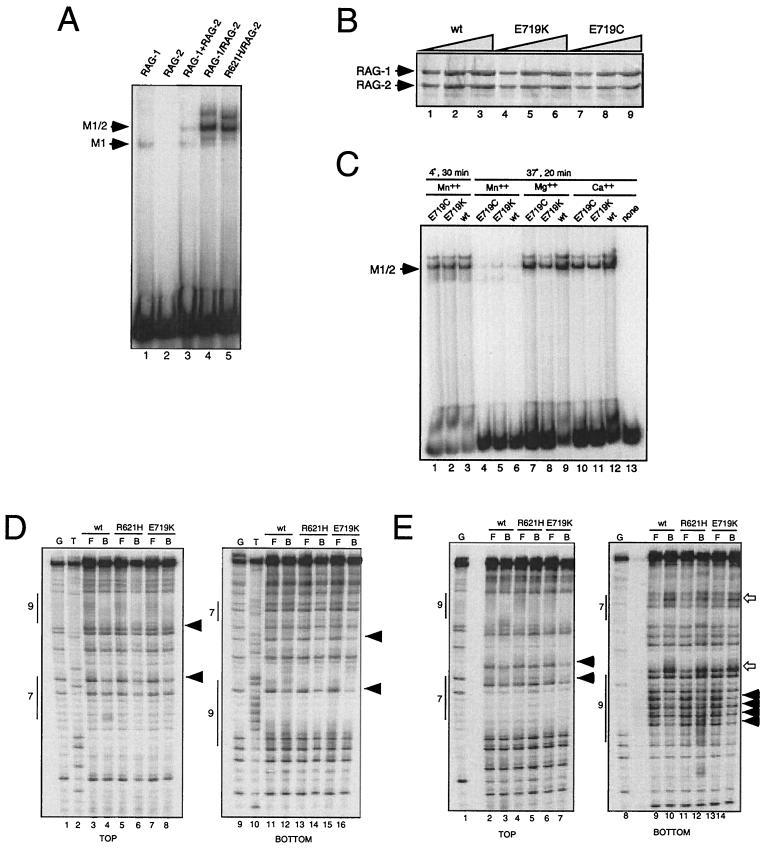
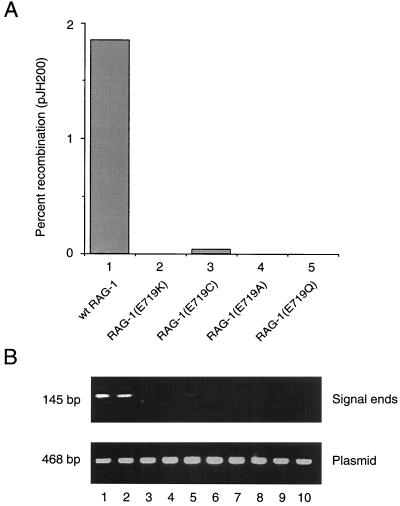
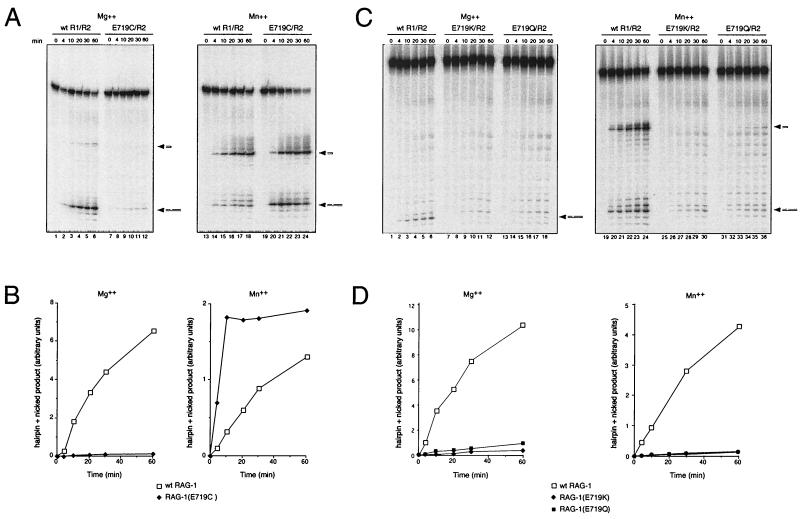
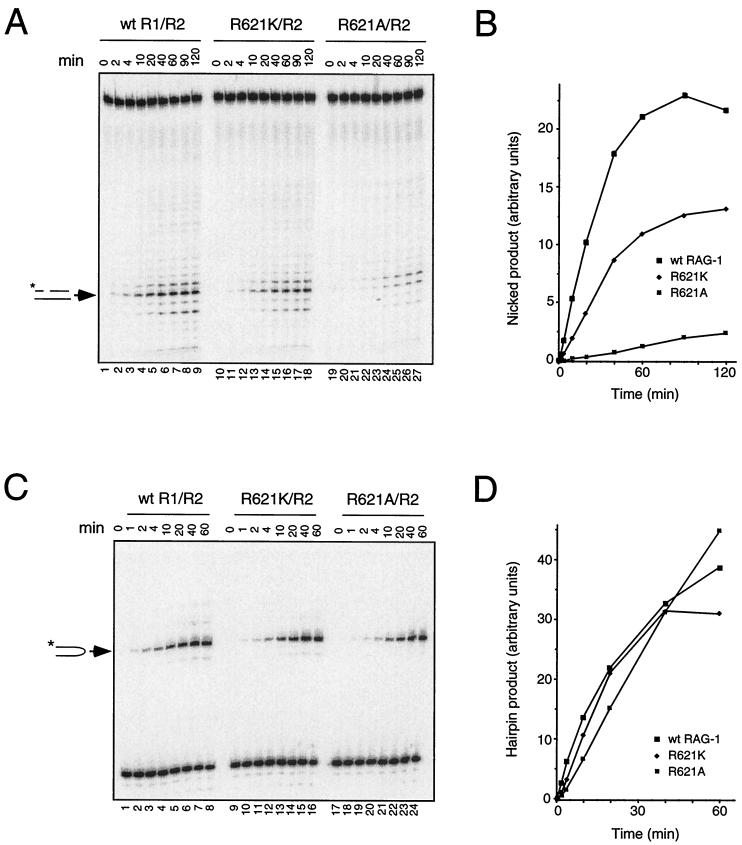
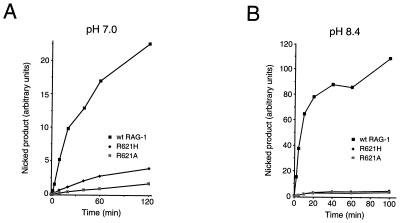
Some patients with B-cell-negative severe combined immune deficiency (SCID) carry mutations in RAG-1 or RAG-2 that impair V(D)J recombination. Two recessive RAG-1 mutations responsible for B-cell-negative SCID, R621H and E719K, impair V(D)J recombination without affecting formation of single-site recombination signal sequence complexes, specific DNA contacts, or perturbation of DNA structure at the heptamer-coding junction. The E719K mutation impairs DNA cleavage by the RAG complex, with a greater effect on nicking than on transesterification; a conservative glutamine substitution exhibits a similar effect. When cysteine is substituted for E719, RAG-1 activity is enhanced in Mn(2+) but remains impaired in Mg(2+), suggesting an interaction between this residue and an essential metal ion. The R621H mutation partially impairs nicking, with little effect on transesterification. The residual nicking activity of the R621H mutant is reduced at least 10-fold upon a change from pH 7.0 to pH 8.4. Site-specific nicking is severely impaired by an alanine substitution at R621 but is spared by substitution with lysine. These observations are consistent with involvement of a positively charged residue at position 621 in the nicking step of the RAG-mediated cleavage reaction. Our data provide a mechanistic explanation for one form of hereditary SCID. Moreover, while RAG-1 is directly involved in catalysis of both nicking and transesterification, our observations indicate that these two steps have distinct catalytic requirements.
Figures
References
-
- Agrawal A, Eastman Q M, Schatz D G. Transposition mediated by RAG1 and RAG2 and its implications for the evolution of the immune system. Nature. 1998;394:744–751. - PubMed
-
- Bolland S, Kleckner N. The three chemical steps of Tn10/IS10 transposition involve repeated utilization of a single active site. Cell. 1996;84:223–233. - PubMed
-
- Bujacz G, Jaskolski M, Alexandratos J, Wlodawer A, Merkel G, Katz R A, Skalka A M. The catalytic domain of avian sarcoma virus integrase: conformation of the active-site residues in the presence of divalent cations. Structure. 1996;4:89–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources