

Regulation of myosin heavy chain expression during rat skeletal muscle development in vitro
- PMID: 11359938
- PMCID: PMC34600
- DOI: 10.1091/mbc.12.5.1499
Regulation of myosin heavy chain expression during rat skeletal muscle development in vitro
Abstract
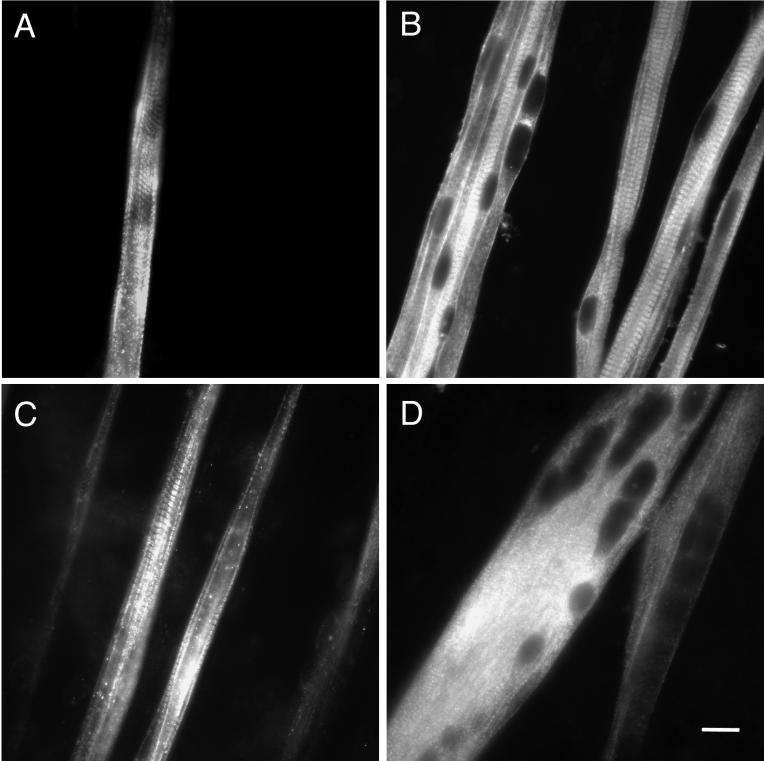
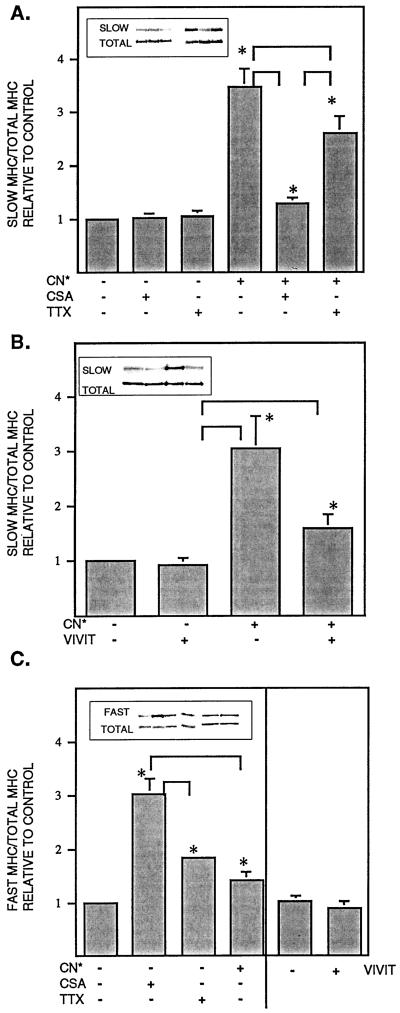
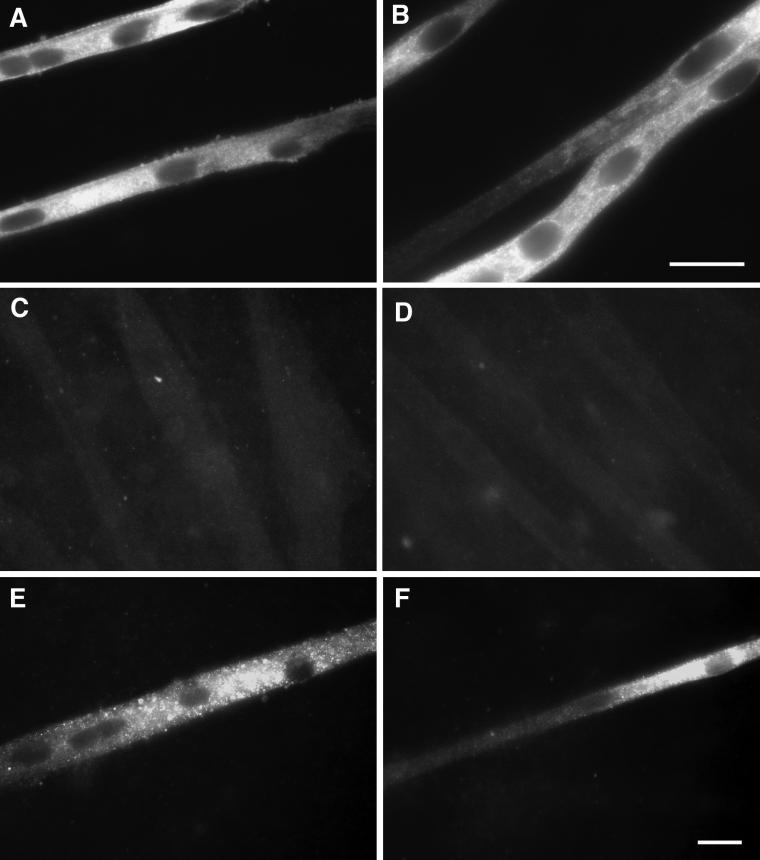
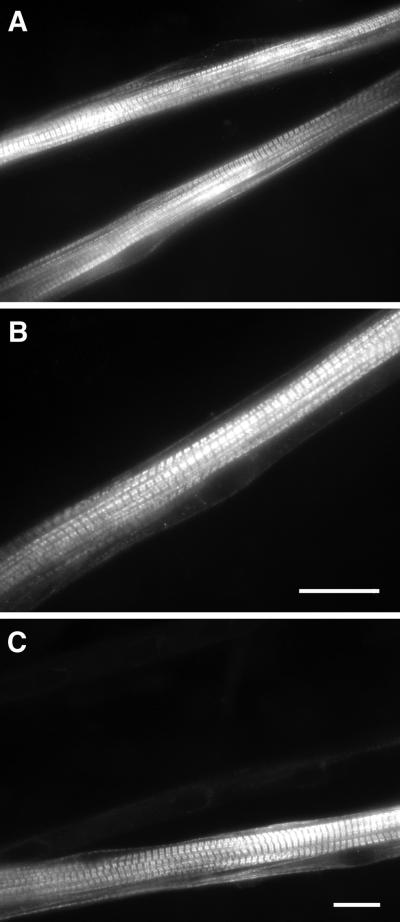
Signals that determine fast- and slow-twitch phenotypes of skeletal muscle fibers are thought to stem from depolarization, with concomitant contraction and activation of calcium-dependent pathways. We examined the roles of contraction and activation of calcineurin (CN) in regulation of slow and fast myosin heavy chain (MHC) protein expression during muscle fiber formation in vitro. Myotubes formed from embryonic day 21 rat myoblasts contracted spontaneously, and approximately 10% expressed slow MHC after 12 d in culture, as seen by immunofluorescent staining. Transfection with a constitutively active form of calcineurin (CN*) increased slow MHC by 2.5-fold as determined by Western blot. This effect was attenuated 35% by treatment with tetrodotoxin and 90% by administration of the selective inhibitor of CN, cyclosporin A. Conversely, cyclosporin A alone increased fast MHC by twofold. Cotransfection with VIVIT, a peptide that selectively inhibits calcineurin-induced activation of the nuclear factor of activated T-cells, blocked the effect of CN* on slow MHC by 70% but had no effect on fast MHC. The results suggest that contractile activity-dependent expression of slow MHC is mediated largely through the CN-nuclear factor of activated T-cells pathway, whereas suppression of fast MHC expression may be independent of nuclear factor of activated T-cells.
Figures
Similar articles
-
Calcineurin regulates slow myosin, but not fast myosin or metabolic enzymes, during fast-to-slow transformation in rabbit skeletal muscle cell culture.J Physiol. 2001 May 15;533(Pt 1):215-26. doi: 10.1111/j.1469-7793.2001.0215b.x. J Physiol. 2001. PMID: 11351029 Free PMC article.
-
The calcineurin-NFAT pathway and muscle fiber-type gene expression.Am J Physiol Cell Physiol. 2000 Oct;279(4):C915-24. doi: 10.1152/ajpcell.2000.279.4.C915. Am J Physiol Cell Physiol. 2000. PMID: 11003571
-
Calcineurin plays a modulatory role in loading-induced regulation of type I myosin heavy chain gene expression in slow skeletal muscle.Am J Physiol Regul Integr Comp Physiol. 2009 Oct;297(4):R1037-48. doi: 10.1152/ajpregu.00349.2009. Epub 2009 Aug 5. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19657098 Free PMC article.
-
Fibre types in skeletal muscle: a personal account.Acta Physiol (Oxf). 2010 Aug;199(4):451-63. doi: 10.1111/j.1748-1716.2010.02130.x. Epub 2010 Mar 26. Acta Physiol (Oxf). 2010. PMID: 20353491 Review.
-
Calcineurin signaling and neural control of skeletal muscle fiber type and size.Trends Pharmacol Sci. 2002 Dec;23(12):569-75. doi: 10.1016/s0165-6147(02)02111-9. Trends Pharmacol Sci. 2002. PMID: 12457775 Review.
Cited by
-
Altered skeletal muscle phenotypes in calcineurin Aalpha and Abeta gene-targeted mice.Mol Cell Biol. 2003 Jun;23(12):4331-43. doi: 10.1128/MCB.23.12.4331-4343.2003. Mol Cell Biol. 2003. PMID: 12773574 Free PMC article.
-
Enhancement of calcium signalling dynamics and stability by delayed modulation of the plasma-membrane calcium-ATPase in human T cells.J Physiol. 2002 Jun 15;541(Pt 3):877-94. doi: 10.1113/jphysiol.2001.016154. J Physiol. 2002. PMID: 12068047 Free PMC article.
-
Self-Healing Scaffolding Technology with Strong, Reversible Interactions under Physiological Conditions for Engineering Marbled Cultured Meat.ACS Appl Mater Interfaces. 2025 Jun 4;17(22):31881-31897. doi: 10.1021/acsami.5c03479. Epub 2025 May 3. ACS Appl Mater Interfaces. 2025. PMID: 40317268 Free PMC article.
-
Spatial Geometries of Self-Assembled Chitohexaose Monolayers Regulate Myoblast Fusion.Int J Mol Sci. 2016 May 6;17(5):686. doi: 10.3390/ijms17050686. Int J Mol Sci. 2016. PMID: 27164094 Free PMC article.
-
Skeletal muscle tissue engineering: a maturation model promoting long-term survival of myotubes, structural development of the excitation-contraction coupling apparatus and neonatal myosin heavy chain expression.Biomaterials. 2009 Oct;30(29):5392-402. doi: 10.1016/j.biomaterials.2009.05.081. Epub 2009 Jul 22. Biomaterials. 2009. PMID: 19625080 Free PMC article.
References
-
- Aramburu J, Yaffe MB, Lopez-Rodriguez C, Cantley LC, Hogan PG, Rao A. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science. 1999;285:2129–2133. - PubMed
-
- Ariano MA, Armstrong RB, Edgerton VR. Hindlimb muscle fiber populations of five mammals. J Histochem Cytochem. 1973;21:51–55. - PubMed
-
- Bigard X, Sanchez H, Zoll J, Mateo P, Rousseau V, Veksler V, Ventura-Clapier R. Calcineurin co-regulates contractile and metabolic components of slow muscle phenotype. J Biol Chem. 2000;275:19653–19660. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
