

Bacillus subtilis locus encoding a killer protein and its antidote
- PMID: 11371520
- PMCID: PMC95233
- DOI: 10.1128/JB.183.12.3574-3581.2001
Bacillus subtilis locus encoding a killer protein and its antidote
Abstract
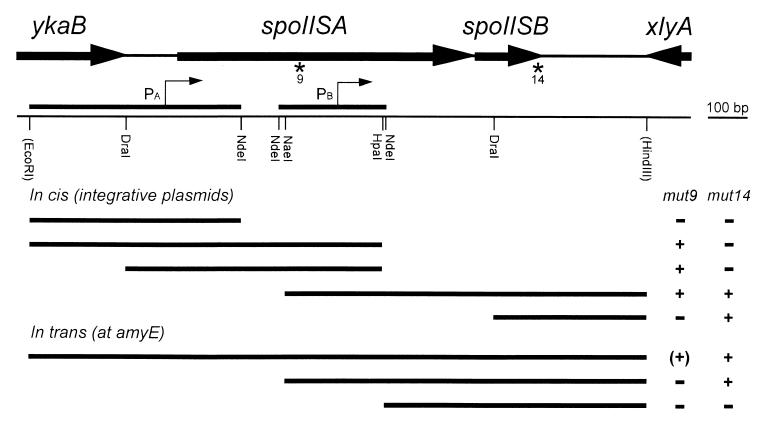
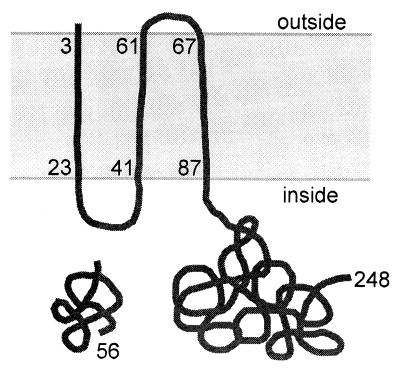
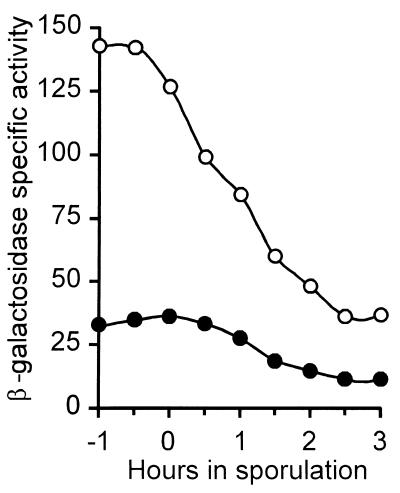
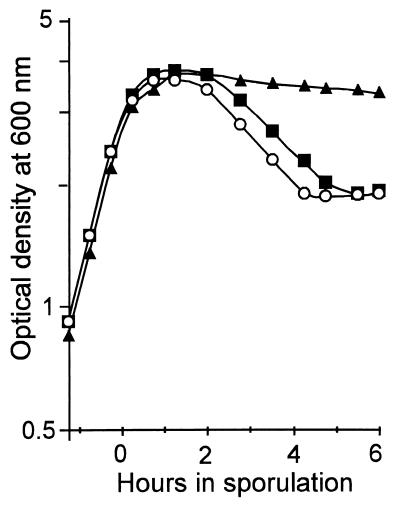
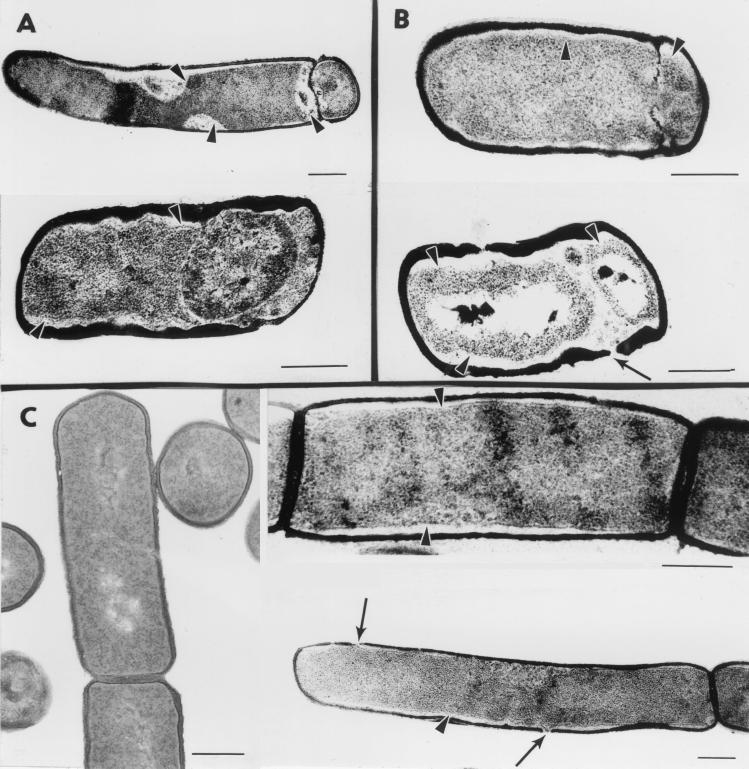
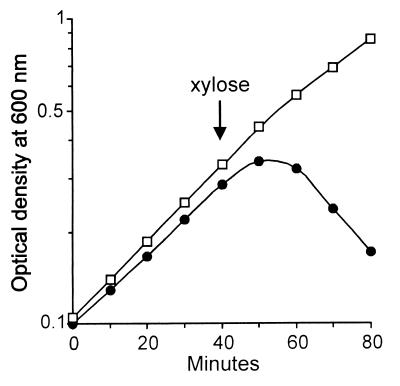
We have isolated mutations that block sporulation after formation of the polar septum in Bacillus subtilis. These mutations were mapped to the two genes of a new locus, spoIIS. Inactivation of the second gene, spoIISB, decreases sporulation efficiency by 4 orders of magnitude. Inactivation of the first gene, spoIISA, has no effect on sporulation but it fully restores sporulation of a spoIISB null mutant, indicating that SpoIISB is required only to counteract the negative effect of SpoIISA on sporulation. An internal promoter ensures the synthesis of an excess of SpoIISB over SpoIISA during exponential growth and sporulation. In the absence of SpoIISB, the sporulating cells show lethal damage of their envelope shortly after asymmetric septation, a defect that can be corrected by synthesizing SpoIISB only in the mother cell. However, forced synthesis of SpoIISA in exponentially growing cells or in the forespore leads to the same type of morphological damage and to cell death. In both cases protection against the killing effect of SpoIISA can be provided by simultaneous synthesis of SpoIISB. The spoIIS locus is unique to B. subtilis, and since it is completely dispensable for sporulation its physiological role remains elusive.
Figures
References
-
- Arigoni F, Guérout-Fleury A-M, Barák I, Stragier P. The SpoIIE phosphatase, the sporulation septum, and the establishment of forespore-specific transcription in Bacillus subtilis: a reassessment. Mol Microbiol. 1999;31:1407–1415. - PubMed
-
- Bai U, Mandic-Mulec I, Smith I. SinI modulates the activity of SinR, a developmental switch protein of Bacillus subtilis, by protein-protein interaction. Genes Dev. 1993;7:139–148. - PubMed
-
- Bishop R E, Leskiw B K, Hodges R S, Kay C M, Weiner J H. The entericidin locus of Escherichia coli and its implications for programmed cell death. J Mol Biol. 1998;280:583–596. - PubMed
-
- Cervin M A, Spiegelman G B, Raether B, Ohlsen K, Perego M, Hoch J A. A negative regulator linking chromosome segregation to developmental transcription in Bacillus subtilis. Mol Microbiol. 1998;29:85–95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
