

Synaptic pattern formation during cellular recognition
- PMID: 11371622
- PMCID: PMC34390
- DOI: 10.1073/pnas.111536798
Synaptic pattern formation during cellular recognition
Abstract
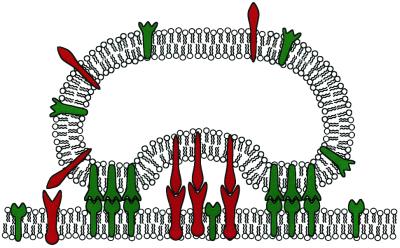
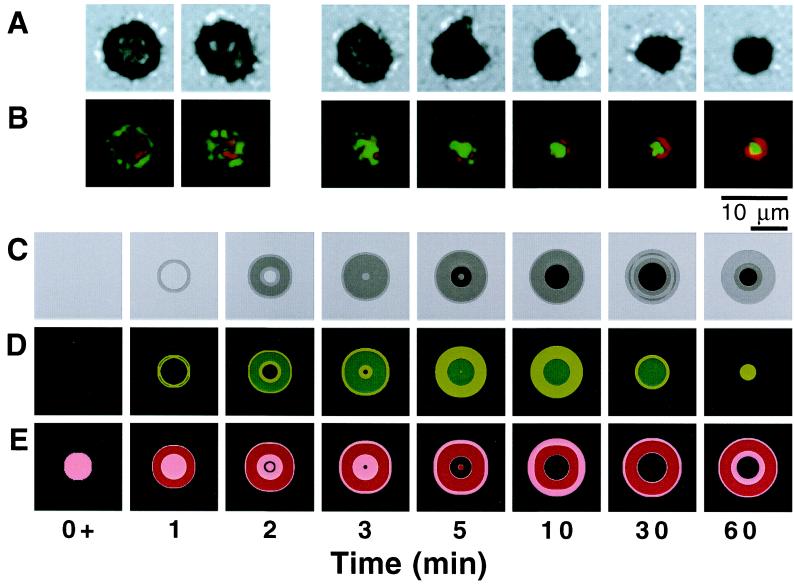
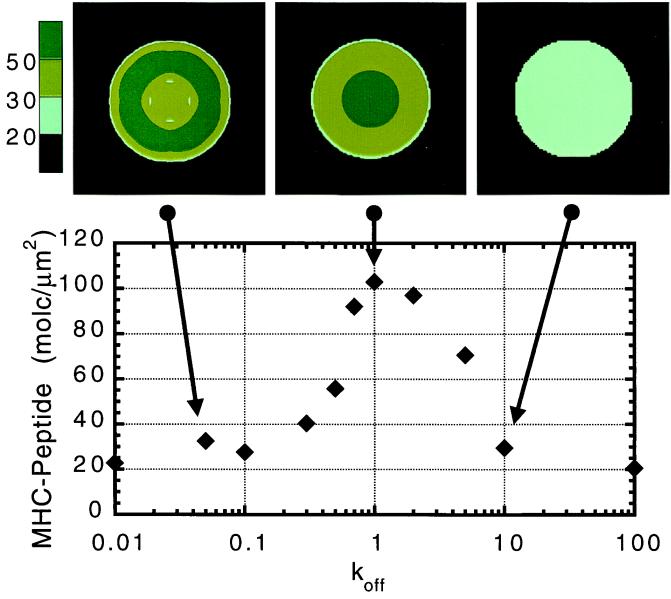
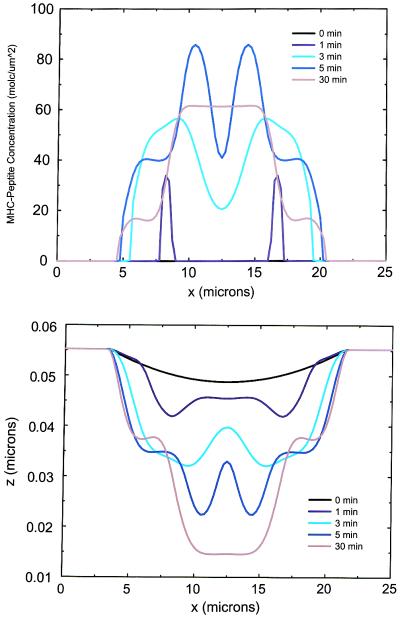
Cell-cell recognition often requires the formation of a highly organized pattern of receptor proteins (a synapse) in the intercellular junction. Recent experiments [e.g., Monks, C. R. F., Freiberg, B. A., Kupfer, H., Sciaky, N. & Kupfer, A. (1998) Nature (London) 395, 82-86; Grakoui, A., Bromley, S. K., Sumen, C., Davis, M. M., Shaw, A. S., Allen, P. M. & Dustin, M. L. (1999) Science 285, 221-227; and Davis, D. M., Chiu, I., Fassett, M., Cohen, G. B., Mandelboim, O. & Strominger, J. L. (1999) Proc. Natl. Acad. Sci. USA 96, 15062-15067] vividly demonstrate a complex evolution of cell shape and spatial receptor-ligand patterns (several microns in size) in the intercellular junction during immunological synapse formation. The current view is that this dynamic rearrangement of proteins into organized supramolecular activation clusters is driven primarily by active cytoskeletal processes [e.g., Dustin, M. L. & Cooper, J. A. (2000) Nat. Immunol. 1, 23-29; and Wulfing, C. & Davis, M. M. (1998) Science 282, 2266-2269]. Here, aided by a quantitative analysis of the relevant physico-chemical processes, we demonstrate that the essential characteristics of synaptic patterns observed in living cells can result from spontaneous self-assembly processes. Active cellular interventions are superimposed on these self-organizing tendencies and may also serve to regulate the spontaneous processes. We find that the protein binding/dissociation characteristics, protein mobilities, and membrane constraints measured in the cellular environment are delicately balanced such that the length and time scales of spontaneously evolving patterns are in near-quantitative agreement with observations for synapse formation between T cells and supported membranes [Grakoui, A., Bromley, S. K., Sumen, C., Davis, M. M., Shaw, A. S., Allen, P. M. & Dustin, M. L. (1999) Science 285, 221-227]. The model we present provides a common way of analyzing immunological synapse formation in disparate systems (e.g., T cell/antigen-presenting cell junctions with different MHC-peptides, natural killer cells, etc.).
Figures
References
-
- Monks C R F, Freiberg B A, Kupfer H, Sciaky N, Kupfer A. Nature (London) 1998;395:82–86. - PubMed
-
- Grakoui A, Bromley S K, Sumen C, Davis M M, Shaw A S, Allen P M, Dustin M L. Science. 1999;285:221–227. - PubMed
-
- Dustin M L, Cooper J A. Nat Immunol. 2000;1:23–29. - PubMed
-
- Wulfing C, Davis M M. Science. 1998;282:2266–2269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
