

The structures of anthranilate synthase of Serratia marcescens crystallized in the presence of (i) its substrates, chorismate and glutamine, and a product, glutamate, and (ii) its end-product inhibitor, L-tryptophan
- PMID: 11371633
- PMCID: PMC33415
- DOI: 10.1073/pnas.111150298
The structures of anthranilate synthase of Serratia marcescens crystallized in the presence of (i) its substrates, chorismate and glutamine, and a product, glutamate, and (ii) its end-product inhibitor, L-tryptophan
Abstract
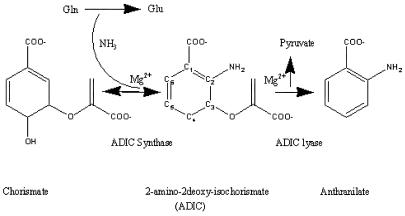
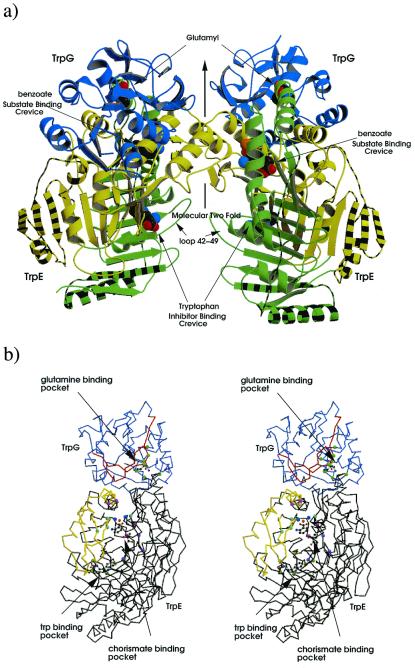
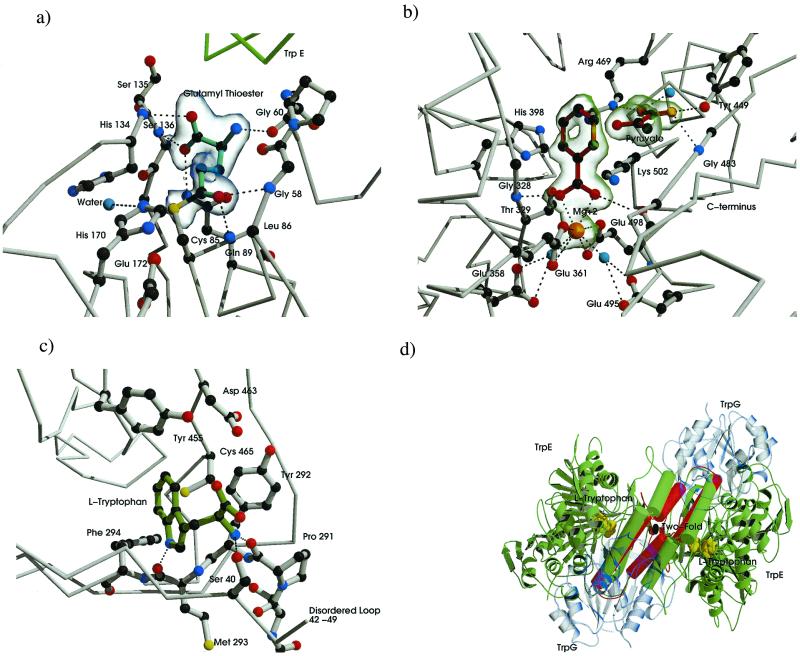
The crystal structure of anthranilate synthase (AS) from Serratia marcescens, a mesophilic bacterium, has been solved in the presence of its substrates, chorismate and glutamine, and one product, glutamate, at 1.95 A, and with its bound feedback inhibitor, tryptophan, at 2.4 A. In comparison with the AS structure from the hyperthermophile Sulfolobus solfataricus, the S. marcescens structure shows similar subunit structures but a markedly different oligomeric organization. One crystal form of the S. marcescens enzyme displays a bound pyruvate as well as a putative anthranilate (the nitrogen group is ambiguous) in the TrpE subunit. It also confirms the presence of a covalently bound glutamyl thioester intermediate in the TrpG subunit. The tryptophan-bound form reveals that the inhibitor binds at a site distinct from that of the substrate, chorismate. Bound tryptophan appears to prevent chorismate binding by a demonstrable conformational effect, and the structure reveals how occupancy of only one of the two feedback inhibition sites can immobilize the catalytic activity of both TrpE subunits. The presence of effectors in the structure provides a view of the locations of some of the amino acid residues in the active sites. Our findings are discussed in terms of the previously described AS structure of S. solfataricus, mutational data obtained from enteric bacteria, and the enzyme's mechanism of action.
Figures

References
-
- Zalkin H, Hwang L H. J Biol Chem. 1971;246:6899–6907. - PubMed
-
- Zalkin H. Adv Enzymol Relat Areas Mol Biol. 1993;66:203–309. - PubMed
-
- Tesmer J J, Klem T J, Deras M L, Davisson V J, Smith J L. Nat Struct Biol. 1996;3:74–86. - PubMed
-
- Bauerle R, Hess J, French S. Methods Enzymol. 1987;142:366–386. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
