

Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity
- PMID: 11371650
- PMCID: PMC33496
- DOI: 10.1073/pnas.101042998
Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity
Abstract
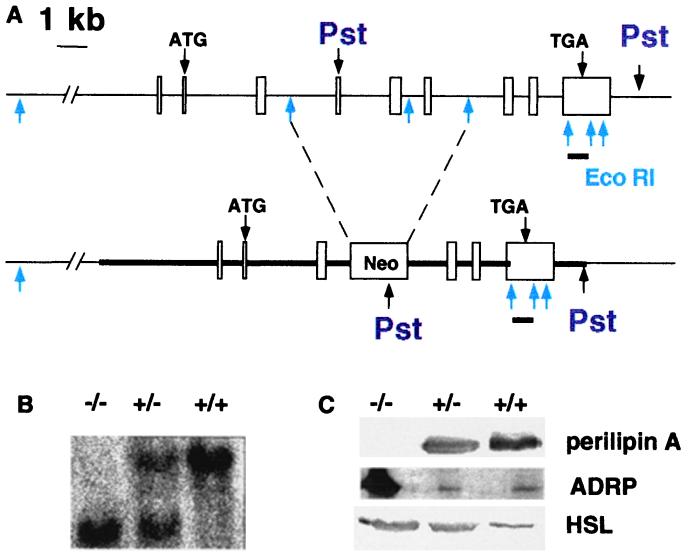
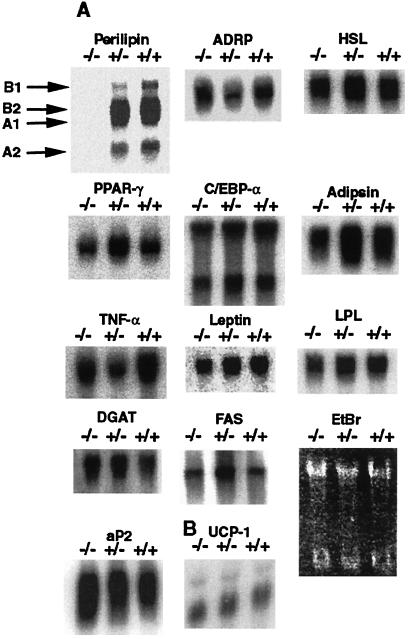
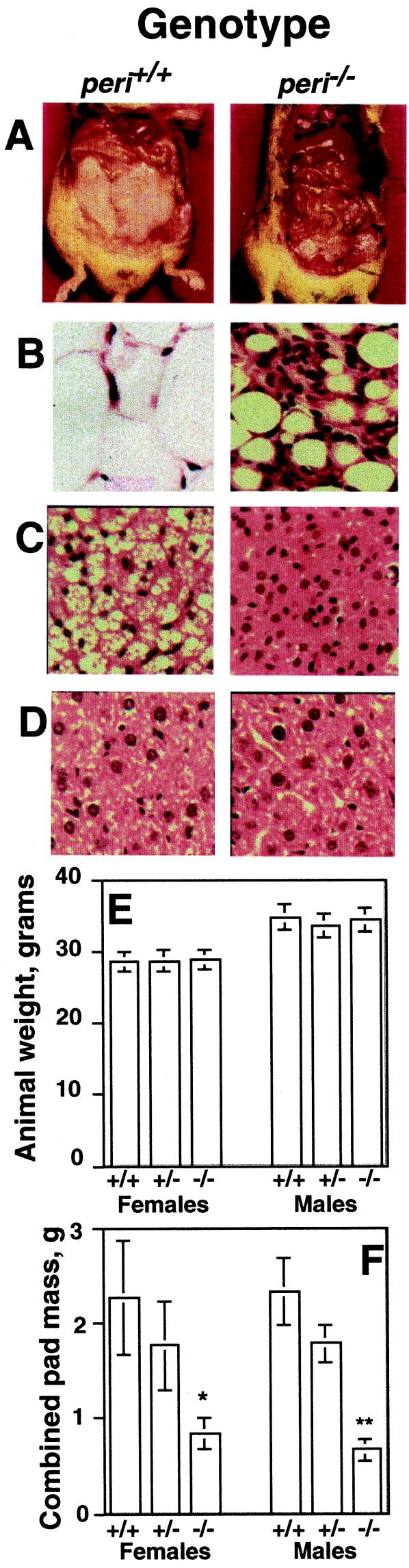
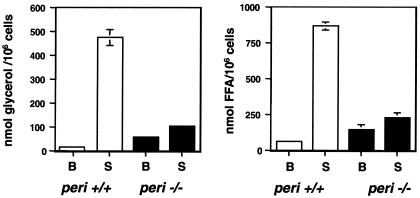
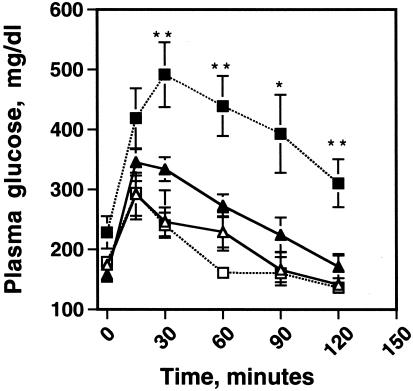
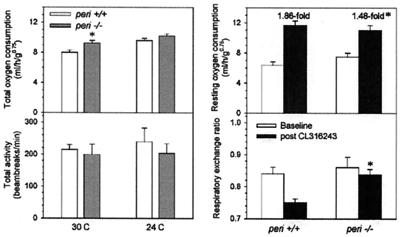
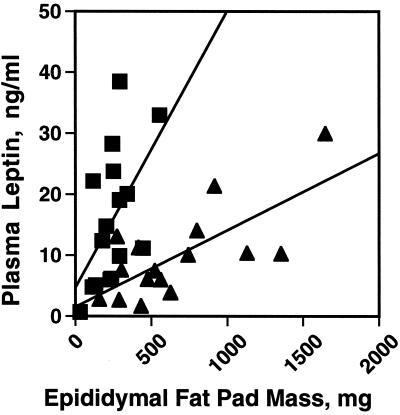
Perilipin coats the lipid droplets of adipocytes and is thought to have a role in regulating triacylglycerol hydrolysis. To study the role of perilipin in vivo, we have created a perilipin knockout mouse. Perilipin null (peri(-/-)) and wild-type (peri(+/+)) mice consume equal amounts of food, but the adipose tissue mass in the null animals is reduced to approximately 30% of that in wild-type animals. Isolated adipocytes of perilipin null mice exhibit elevated basal lipolysis because of the loss of the protective function of perilipin. They also exhibit dramatically attenuated stimulated lipolytic activity, indicating that perilipin is required for maximal lipolytic activity. Plasma leptin concentrations in null animals were greater than expected for the reduced adipose mass. The peri(-/-) animals have a greater lean body mass and increased metabolic rate but they also show an increased tendency to develop glucose intolerance and peripheral insulin resistance. When fed a high-fat diet, the perilipin null animals are resistant to diet-induced obesity but not to glucose intolerance. The data reveal a major role for perilipin in adipose lipid metabolism and suggest perilipin as a potential target for attacking problems associated with obesity.
Figures
References
-
- Servetnick D A, Brasaemle D L, Gruia-Gray J, Kimmel A R, Wolff J, Londos C. J Biol Chem. 1995;270:16970–16973. - PubMed
-
- Londos C, Brasaemle D L, Schultz C J, Segrest J P, Kimmel A R. Semin Cell Dev Biol. 1999;10:51–58. - PubMed
-
- Brasaemle D L, Rubin B, Harten I A, Gruia-Gray J, Kimmel A R, Londos C. J Biol Chem. 2000;275:38486–38493. - PubMed
-
- Souza S C, de Vargas L M, Yamamoto M T, Lien P, Franciosa M D, Moss L G, Greenberg A S. J Biol Chem. 1998;273:24665–24669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
