

Cloning of a phenol oxidase gene from Acremonium murorum and its expression in Aspergillus awamori
- PMID: 11375170
- PMCID: PMC92914
- DOI: 10.1128/AEM.67.6.2610-2616.2001
Cloning of a phenol oxidase gene from Acremonium murorum and its expression in Aspergillus awamori
Abstract
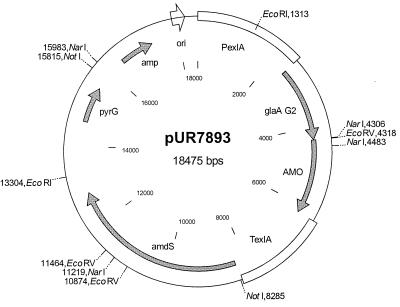
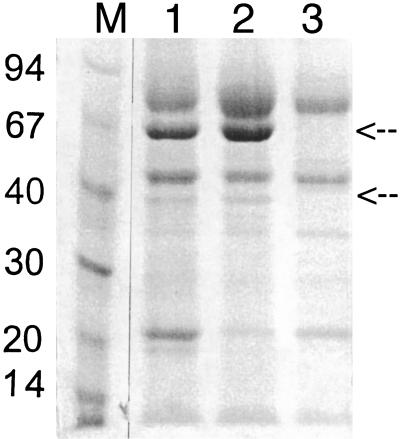
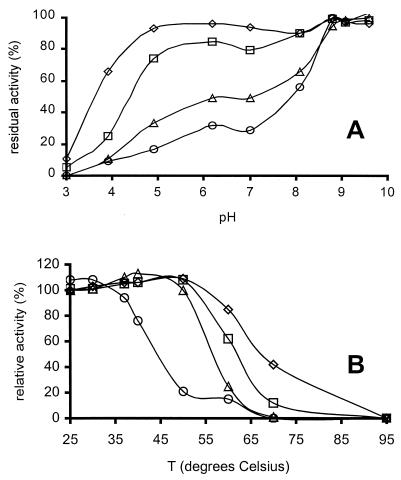
Fungal multicopper oxidases have many potential industrial applications, since they perform reactions under mild conditions. We isolated a phenol oxidase from the fungus Acremonium murorum var. murorum that was capable of decolorizing plant chromophores (such as anthocyanins). This enzyme is of interest in laundry-cleaning products because of its broad specificity for chromophores. We expressed an A. murorum cDNA library in Saccharomyces cerevisiae and subsequently identified enzyme-producing yeast colonies based on their ability to decolor a plant chromophore. The cDNA sequence contained an open reading frame of 1,806 bp encoding an enzyme of 602 amino acids. The phenol oxidase was overproduced by Aspergillus awamori as a fusion protein with glucoamylase, cleaved in vivo, and purified from the culture broth by hydrophobic-interaction chromatography. The phenol oxidase is active at alkaline pH (the optimum for syringaldazine is pH 9) and high temperature (optimum, 60 degrees C) and is fully stable for at least 1 h at 60 degrees C under alkaline conditions. These characteristics and the high production level of 0.6 g of phenol oxidase per liter in shake flasks, which is equimolar with the glucoamylase protein levels, make this enzyme suitable for use in processes that occur under alkaline conditions, such as laundry cleaning.
Figures


References
-
- Bennett J W, Lasure L L. Growth media. In: Bennett J W, Lasure L L, editors. More gene manipulations in fungi. San Diego, Calif: Academic Press, Inc.; 1991. pp. 441–458.
-
- Britton H T S, Robinson R. Universal buffer solutions and the dissociation constant of veronal. J Chem Soc. 1931;458:1456.
-
- Cullen D. Recent advances on the molecular genetics of lignolytic fungi. J Biotechnol. 1997;53:273–289. - PubMed
-
- Gietz R D, Woods R A. Transformation of yeast by the lithium acetate/single stranded carrier DNA/PEG method. Methods Microbiol. 1998;26:53–66.
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
