

Arabidopsis thaliana Rop GTPases are localized to tips of root hairs and control polar growth
- PMID: 11387211
- PMCID: PMC125484
- DOI: 10.1093/emboj/20.11.2779
Arabidopsis thaliana Rop GTPases are localized to tips of root hairs and control polar growth
Abstract
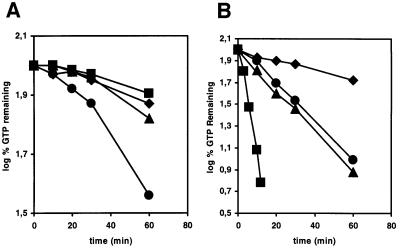
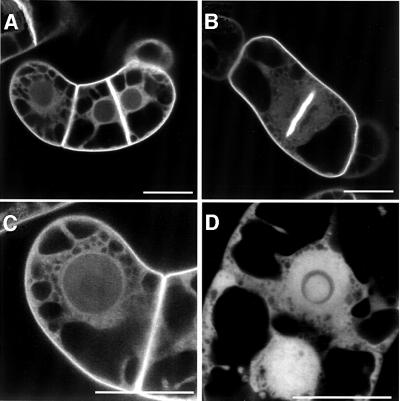
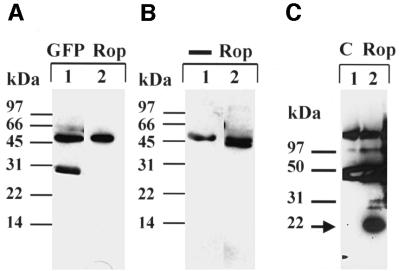
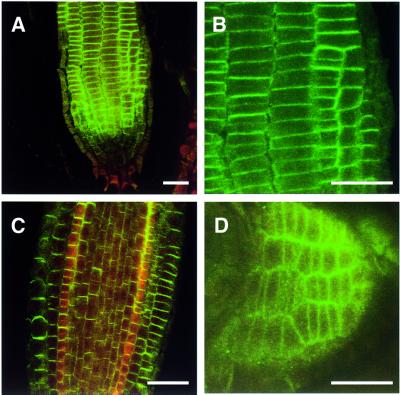
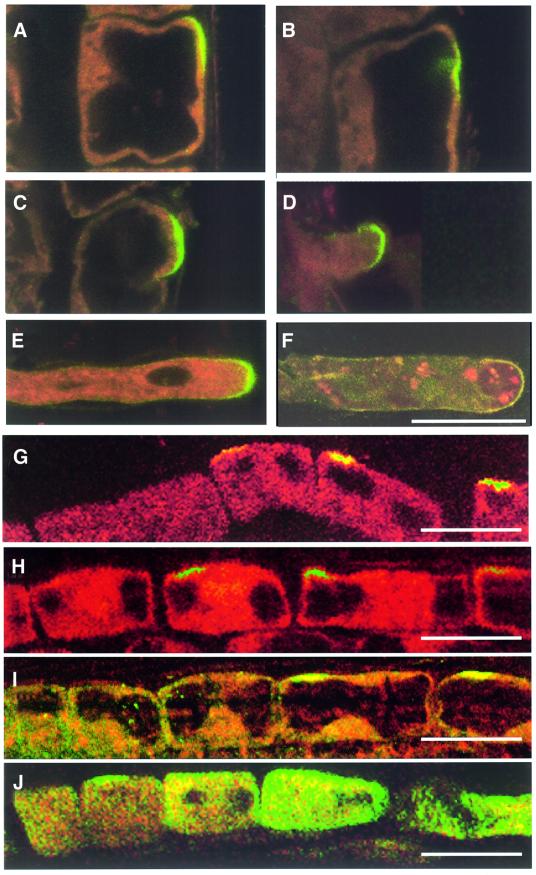
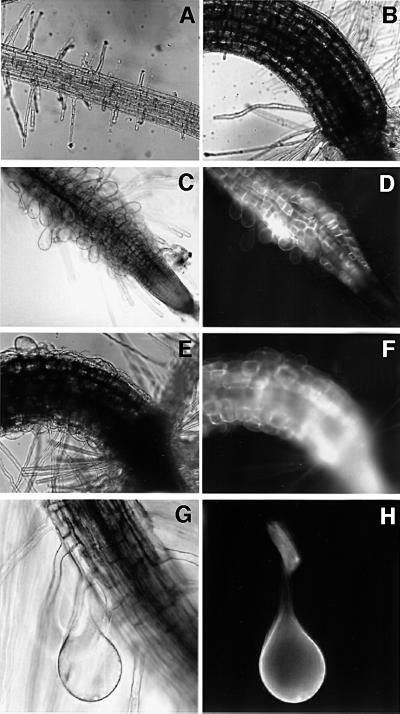
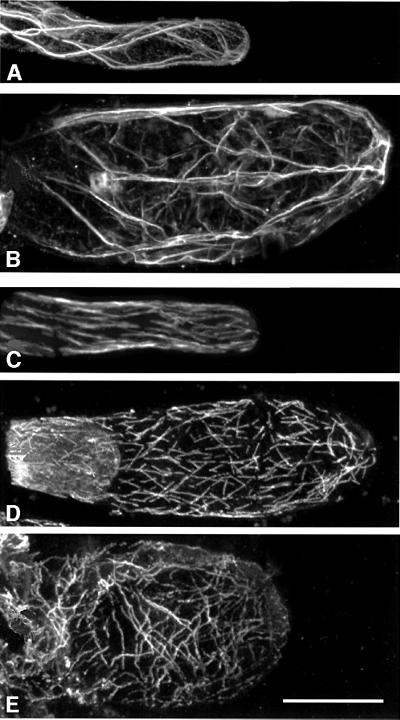
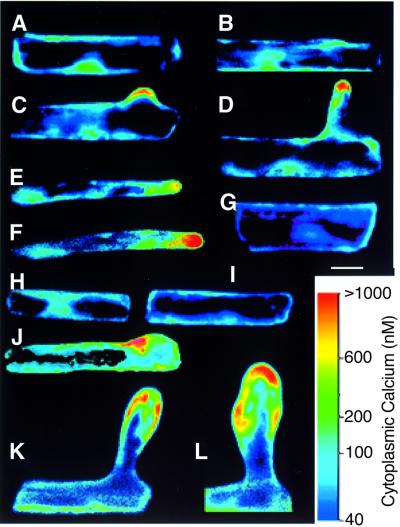
Plants contain a novel unique subfamily of Rho GTPases, vital components of cellular signalling networks. Here we report a general role for some members of this family in polarized plant growth processes. We show that Arabidopsis AtRop4 and AtRop6 encode functional GTPases with similar intrinsic GTP hydrolysis rates. We localized AtRop proteins in root meristem cells to the cross-wall and cell plate membranes. Polar localization of AtRops in trichoblasts specifies the growth sites for emerging root hairs. These sites were visible before budding and elongation of the Arabidopsis root hair when AtRops accumulated at their tips. Expression of constitutively active AtRop4 and AtRop6 mutant proteins in root hairs of transgenic Arabidopsis plants abolished polarized growth and delocalized the tip-focused Ca2+ gradient. Polar localization of AtRops was inhibited by brefeldin A, but not by other drugs such as latrunculin B, cytochalasin D or caffeine. Our results demonstrate a general function of AtRop GTPases in tip growth and in polar diffuse growth.
Figures

Similar articles
-
The Arabidopsis Rab GTPase RabA4b localizes to the tips of growing root hair cells.Plant Cell. 2004 Jun;16(6):1589-603. doi: 10.1105/tpc.021634. Epub 2004 May 21. Plant Cell. 2004. PMID: 15155878 Free PMC article.
-
Co-regulation of root hair tip growth by ROP GTPases and nitrogen source modulated pH fluctuations.Plant Signal Behav. 2011 Mar;6(3):426-9. doi: 10.4161/psb.6.3.14523. Plant Signal Behav. 2011. PMID: 21673509 Free PMC article.
-
Localization of AtROP4 and AtROP6 and interaction with the guanine nucleotide dissociation inhibitor AtRhoGDI1 from Arabidopsis.Plant Mol Biol. 2000 Feb;42(3):515-30. doi: 10.1023/a:1006341210147. Plant Mol Biol. 2000. PMID: 10798620
-
Building a hair: tip growth in Arabidopsis thaliana root hairs.Philos Trans R Soc Lond B Biol Sci. 2002 Jun 29;357(1422):815-21. doi: 10.1098/rstb.2002.1092. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12079677 Free PMC article. Review.
-
Vesicular trafficking, cytoskeleton and signalling in root hairs and pollen tubes.Trends Plant Sci. 2006 Dec;11(12):594-600. doi: 10.1016/j.tplants.2006.10.002. Epub 2006 Nov 7. Trends Plant Sci. 2006. PMID: 17092761 Review.
Cited by
-
Microtubule-associated ROP interactors affect microtubule dynamics and modulate cell wall patterning and root hair growth.Development. 2022 Nov 15;149(22):dev200811. doi: 10.1242/dev.200811. Epub 2022 Nov 16. Development. 2022. PMID: 36314989 Free PMC article.
-
A comparative analysis of proteins that accumulate during the initial stage of root hair development in barley root hair mutants and their parent varieties.J Appl Genet. 2012 Nov;53(4):363-76. doi: 10.1007/s13353-012-0105-1. Epub 2012 Jul 31. J Appl Genet. 2012. PMID: 22847350 Free PMC article.
-
Root hairs.Arabidopsis Book. 2002;1:e0060. doi: 10.1199/tab.0060. Epub 2002 Apr 4. Arabidopsis Book. 2002. PMID: 22303213 Free PMC article. No abstract available.
-
Analysis of the small GTPase gene superfamily of Arabidopsis.Plant Physiol. 2003 Mar;131(3):1191-208. doi: 10.1104/pp.013052. Plant Physiol. 2003. PMID: 12644670 Free PMC article.
-
RAPTOR Controls Developmental Growth Transitions by Altering the Hormonal and Metabolic Balance.Plant Physiol. 2018 Jun;177(2):565-593. doi: 10.1104/pp.17.01711. Epub 2018 Apr 23. Plant Physiol. 2018. PMID: 29686055 Free PMC article.
References
-
- Aoyama T. and Chua,N.-H. (1997) A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J., 11, 605–612. - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature, 408, 796–815. - PubMed
-
- Baluška F., Barlow,P.W. and Volkmann,D. (2000a) Actin and myosin VII in developing root cells. In Staiger,C.J., Baluška,F., Volkmann,D. and Barlow,P.W. (eds), Actin: A Dynamic Framework for Multiple Plant Cell Function. Kluwer Academic, Dordrecht, The Netherlands, pp. 457–476.
-
- Baluška F., Salaj,J., Mathur,J., Braun,M., Jasper,F., Šamaj,J., Chua, N.-H., Barlow,P.W. and Volkmann,D. (2000b) Root hair formation: F-actin dependent tip growth is initiated by a local assembly of profilin-supported F-actin meshworks accumulated within expansin-enriched bulges. Dev. Biol., 227, 618–632. - PubMed
-
- Barlow P.W. and Baluška,F. (2000) Cytoskeletal perspectives on root growth and morphogenesis. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51, 289–322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
