

Requirement of Hsp90 for centrosomal function reflects its regulation of Polo kinase stability
- PMID: 11387220
- PMCID: PMC125474
- DOI: 10.1093/emboj/20.11.2878
Requirement of Hsp90 for centrosomal function reflects its regulation of Polo kinase stability
Abstract
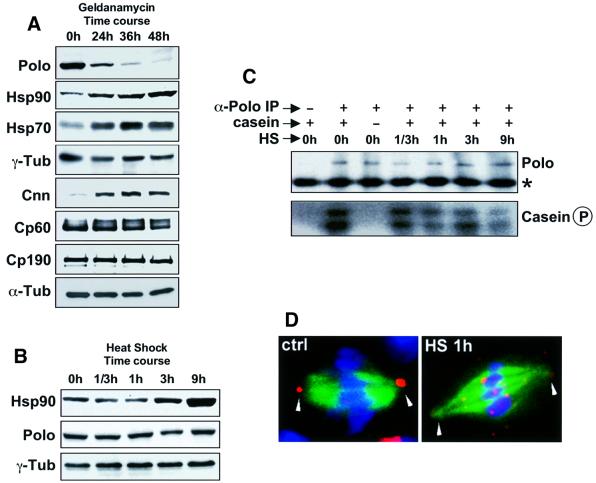
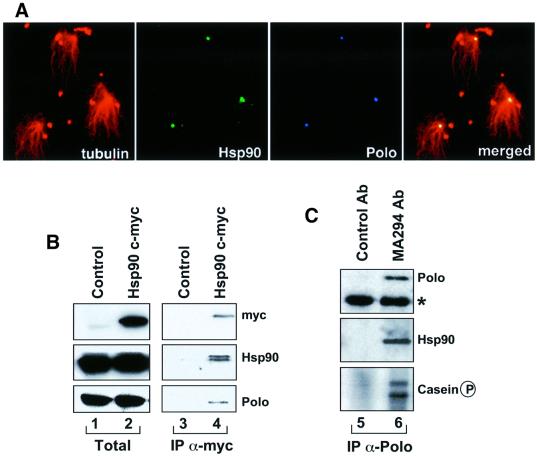
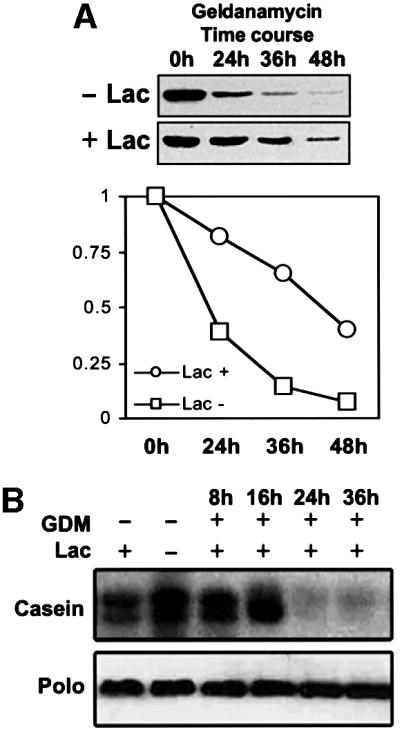
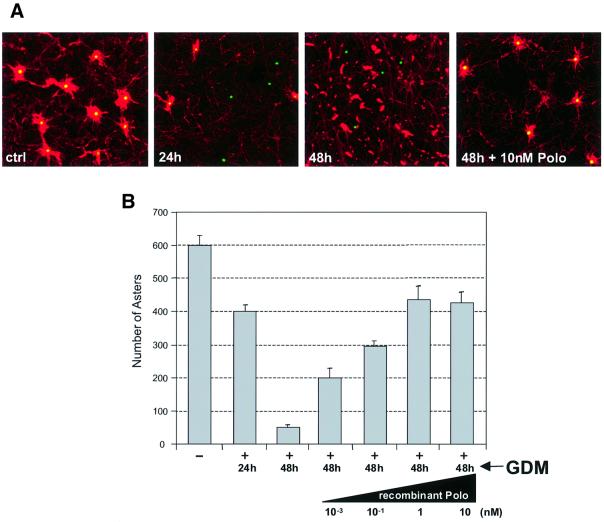
We have previously shown that the molecular chaperone heat shock protein 90 (Hsp90) is required to ensure proper centrosome function in Drosophila and vertebrate cells. This observation led to the hypothesis that this chaperone could be required for the stability of one or more centrosomal proteins. We have found that one of these is Polo, a protein kinase known to regulate several aspects of cell division including centrosome maturation and function. Inhibition of Hsp90 results in the inactivation of Polo kinase activity. It also leads to a loss in the ability of cytoplasmic extracts to complement the failure of salt-stripped preparations of centrosomes to nucleate microtubules. This effect can be rescued upon addition of active recombinant POLO: We also show that Polo and Hsp90 are part of a complex and conclude that stabilization of Polo is one of the mechanisms by which Hsp90 contributes to the maintenance of functional centrosomes.
Figures
References
-
- Avides M.C. and Glover,D.M. (1999) Abnormal spindle protein, Asp and the integrity of mitotic centrosomal microtubule organizing centres. Science, 283, 1733–1735. - PubMed
-
- Barnett C.A., Palmour,R.M., Litwack,G. and Seegmiller,J.E. (1983) In vitro stabilization of the unoccupied glucocorticoid receptor by adenosine 5′-diphosphate. Endocrinology, 112, 2059–2068. - PubMed
-
- Baulieu E.E. and Jung,I. (1972) Aprostatic cytosol ‘receptor’ in the rat levator ani muscle. Nature New Biol., 237, 24–26. - PubMed
-
- Buchner J. (1999) Hsp90 & Co. a holding for folding. Trends Biochem. Sci., 24, 136–141. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
