

Altered nucleotide misinsertion fidelity associated with poliota-dependent replication at the end of a DNA template
- PMID: 11387224
- PMCID: PMC125476
- DOI: 10.1093/emboj/20.11.2914
Altered nucleotide misinsertion fidelity associated with poliota-dependent replication at the end of a DNA template
Abstract
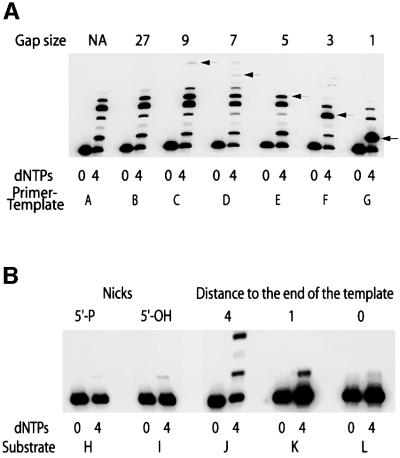
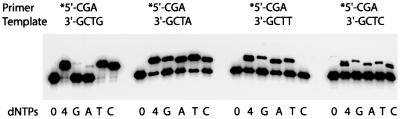
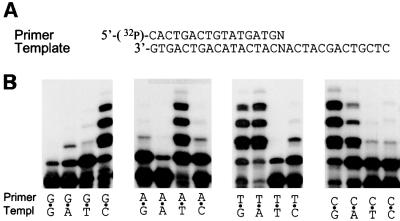
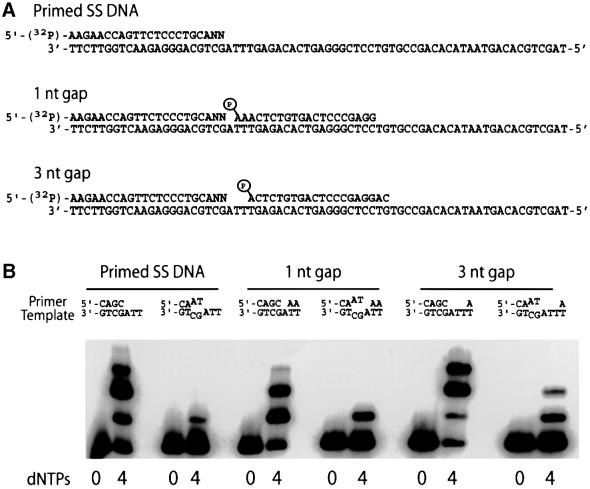
A hallmark of human DNA polymerase iota (poliota) is the asymmetric fidelity of replication at template A and T when the enzyme extends primers annealed to a single-stranded template. Here, we report on the efficiency and accuracy of poliota-dependent replication at a nick, a gap, the very end of a template and from a mispaired primer. Poliota cannot initiate synthesis on a nicked DNA substrate, but fills short gaps efficiently. Surprisingly, poliota's ability to blunt-end a 1 bp recessed terminus is dependent upon the template nucleotide encountered and is highly erroneous. At template G, both C and T are inserted with roughly equal efficiency, whilst at template C, C and A are misinserted 8- and 3-fold more often than the correct base, G. Using substrates containing mispaired primer termini, we show that poliota can extend all 12 mispairs, but with differing efficiencies. Poliota can also extend a tandem mispair, especially when it is located within a short gap. The enzymatic properties of poliota appear consistent with that of a somatic hypermutase and suggest that poliota may be one of the low-fidelity DNA polymerases hypothesized to participate in the hypermutation of immunoglobulin variable genes in vivo.
Figures

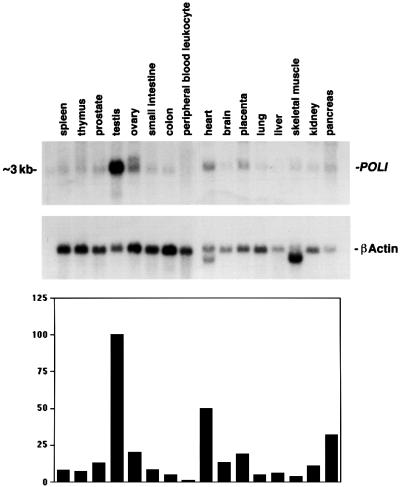
References
-
- Bebenek K., Tissier,A., Frank,E.G., McDonald,J.P., Prasad,R., Wilson,S.H., Woodgate,R. and Kunkel,T.A. (2001) 5′-deoxyribose phosphate lyase activity of human DNA polymerase ι in vitro. Science, 291, 2156–2159. - PubMed
-
- Betz A.G., Milstein,C., Gonzalez-Fernandez,A., Pannell,R., Larson,T. and Neuberger,M.S. (1994) Elements regulating somatic hypermutation of an immunoglobulin κ gene: critical role for the intron enhancer/matrix attachment region. Cell, 77, 239–248. - PubMed
-
- Boosalis M.S., Petruska,J. and Goodman,M.F. (1987) DNA polymerase insertion fidelity. Gel assay for site-specific kinetics. J. Biol. Chem., 262, 14689–14696. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
