

Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast
- PMID: 11390663
- PMCID: PMC87095
- DOI: 10.1128/MCB.21.13.4347-4368.2001
Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast
Abstract
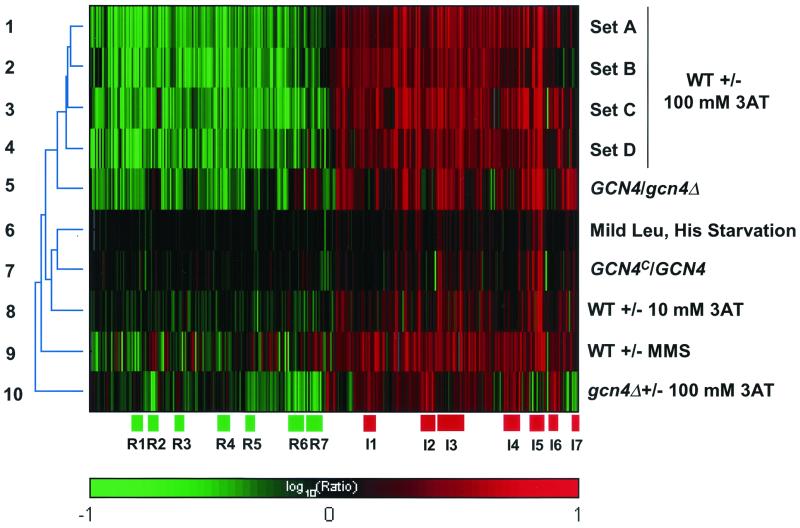
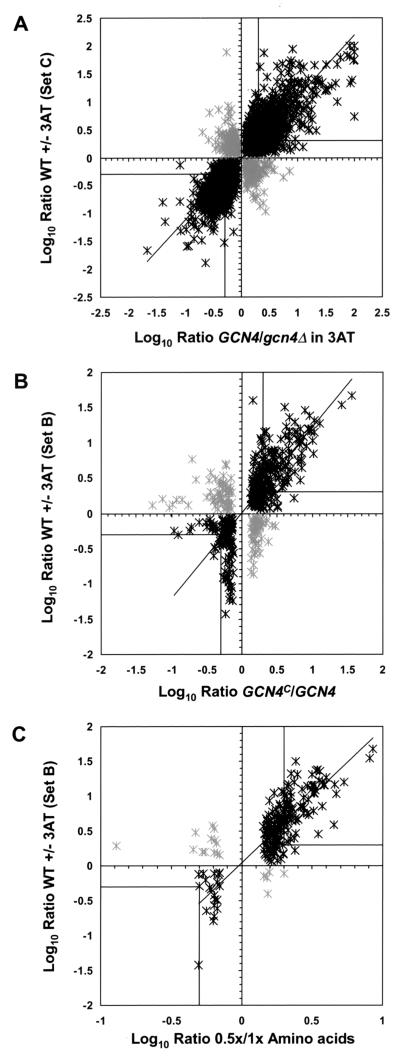
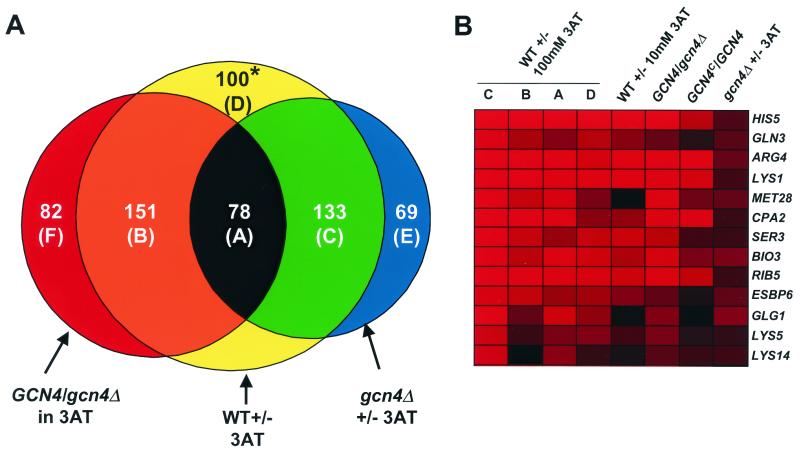
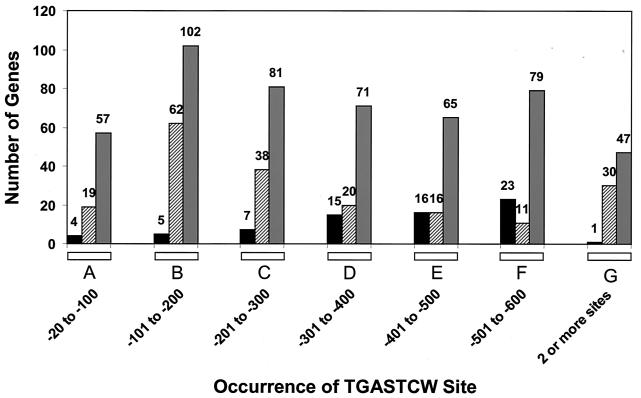
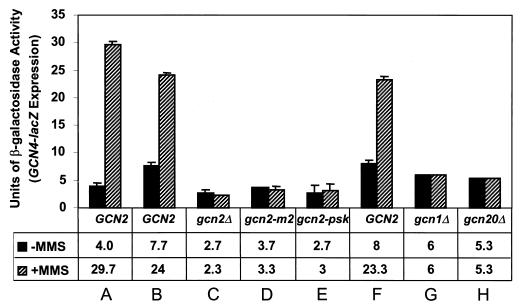
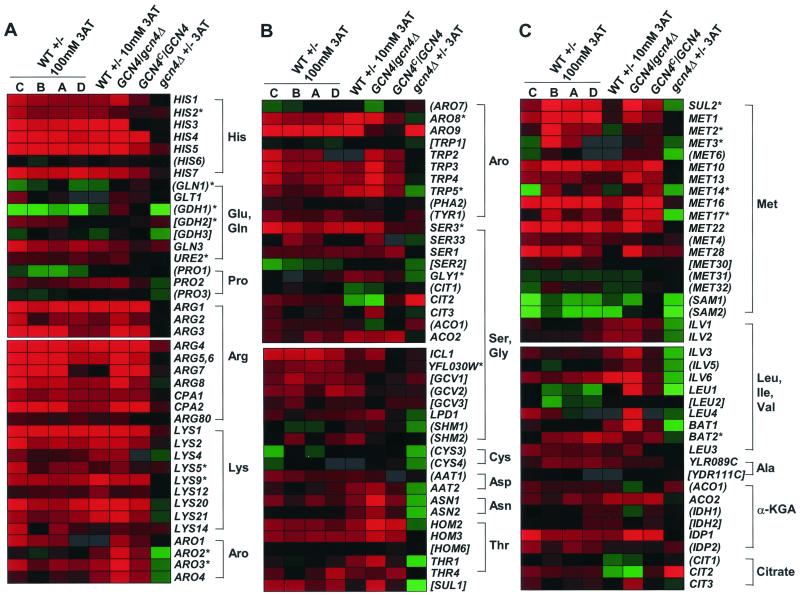
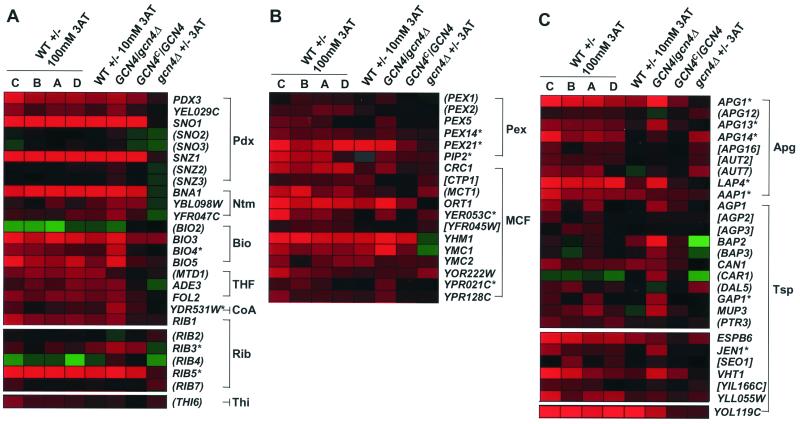
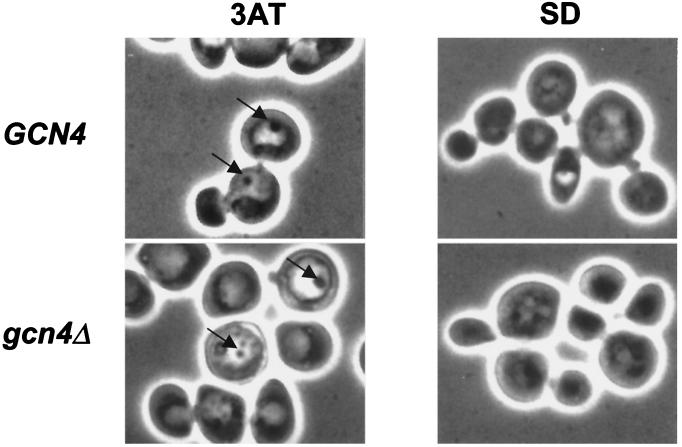
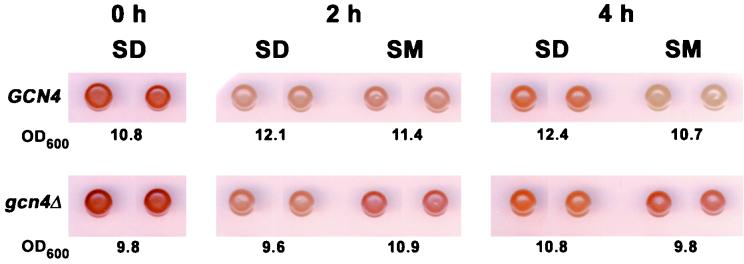
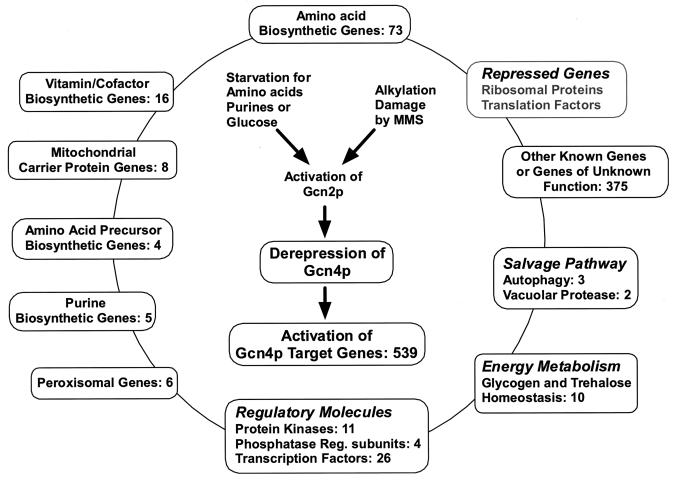
Starvation for amino acids induces Gcn4p, a transcriptional activator of amino acid biosynthetic genes in Saccharomyces cerevisiae. In an effort to identify all genes regulated by Gcn4p during amino acid starvation, we performed cDNA microarray analysis. Data from 21 pairs of hybridization experiments using two different strains derived from S288c revealed that more than 1,000 genes were induced, and a similar number were repressed, by a factor of 2 or more in response to histidine starvation imposed by 3-aminotriazole (3AT). Profiling of a gcn4Delta strain and a constitutively induced mutant showed that Gcn4p is required for the full induction by 3AT of at least 539 genes, termed Gcn4p targets. Genes in every amino acid biosynthetic pathway except cysteine and genes encoding amino acid precursors, vitamin biosynthetic enzymes, peroxisomal components, mitochondrial carrier proteins, and autophagy proteins were all identified as Gcn4p targets. Unexpectedly, genes involved in amino acid biosynthesis represent only a quarter of the Gcn4p target genes. Gcn4p also activates genes involved in glycogen homeostasis, and mutant analysis showed that Gcn4p suppresses glycogen levels in amino acid-starved cells. Numerous genes encoding protein kinases and transcription factors were identified as targets, suggesting that Gcn4p is a master regulator of gene expression. Interestingly, expression profiles for 3AT and the alkylating agent methyl methanesulfonate (MMS) overlapped extensively, and MMS induced GCN4 translation. Thus, the broad transcriptional response evoked by Gcn4p is produced by diverse stress conditions. Finally, profiling of a gcn4Delta mutant uncovered an alternative induction pathway operating at many Gcn4p target genes in histidine-starved cells.
Figures
References
-
- Albrecht G, Mosch H U, Hoffmann B, Reusser U, Braus G H. Monitoring the Gcn4 protein-mediated response in the yeast Saccharomyces cerevisiae. J Biol Chem. 1998;273:12696–12702. - PubMed
-
- Arndt K T, Styles C, Fink G R. Multiple global regulators control HIS4 transcription in yeast. Science. 1987;237:874–880. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases