

Complex functions of AP-1 transcription factors in differentiation and survival of PC12 cells
- PMID: 11390664
- PMCID: PMC87096
- DOI: 10.1128/MCB.21.13.4369-4378.2001
Complex functions of AP-1 transcription factors in differentiation and survival of PC12 cells
Abstract
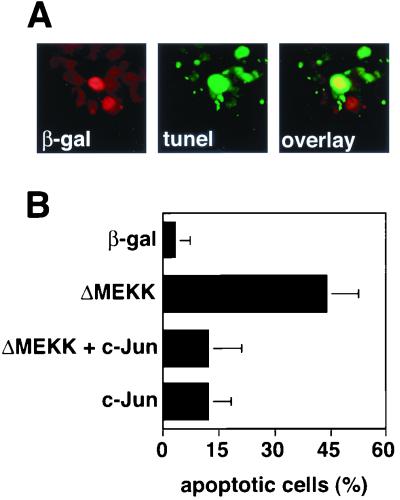
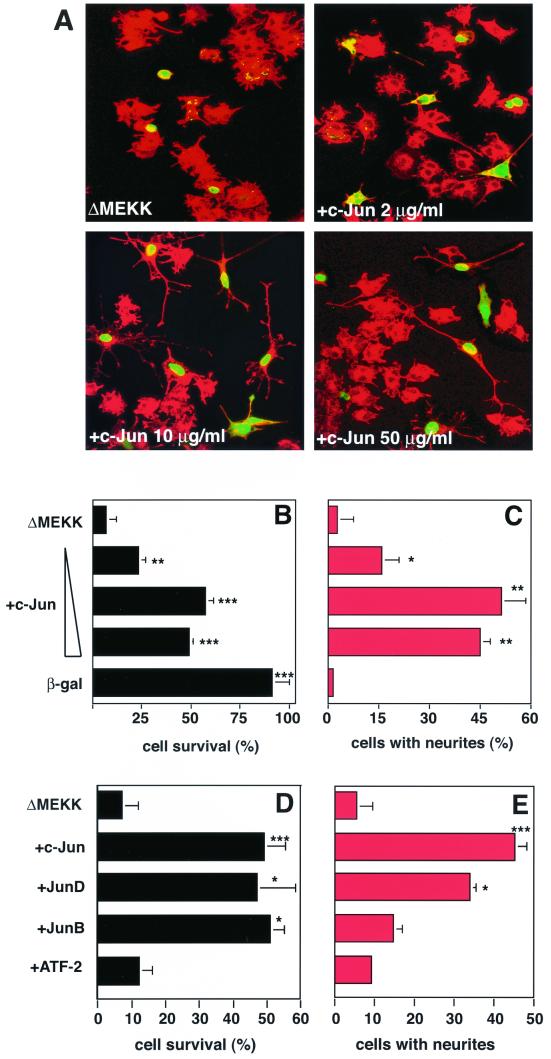
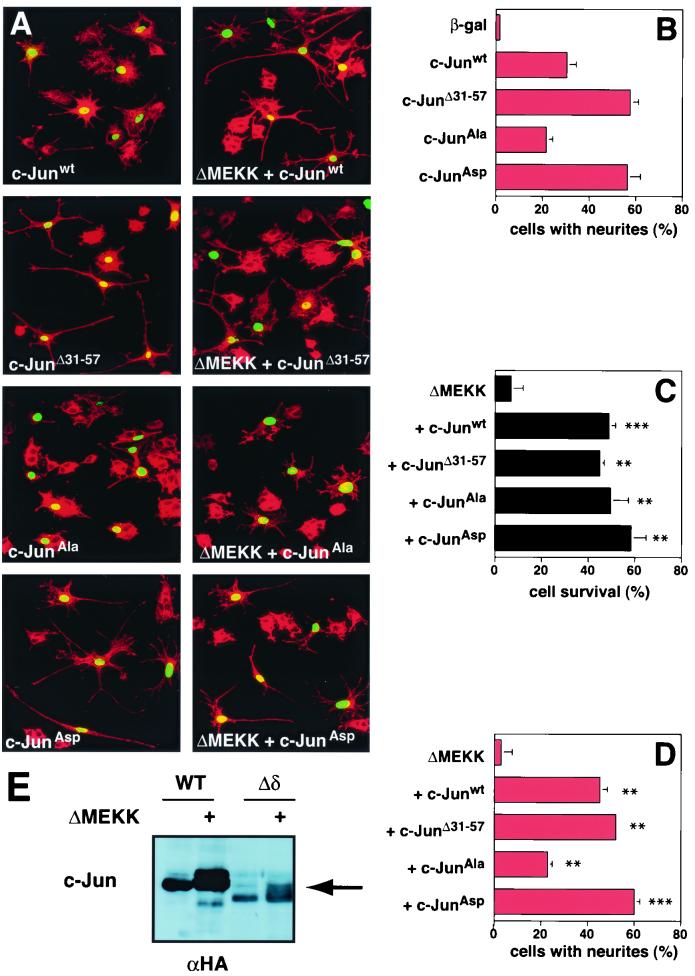
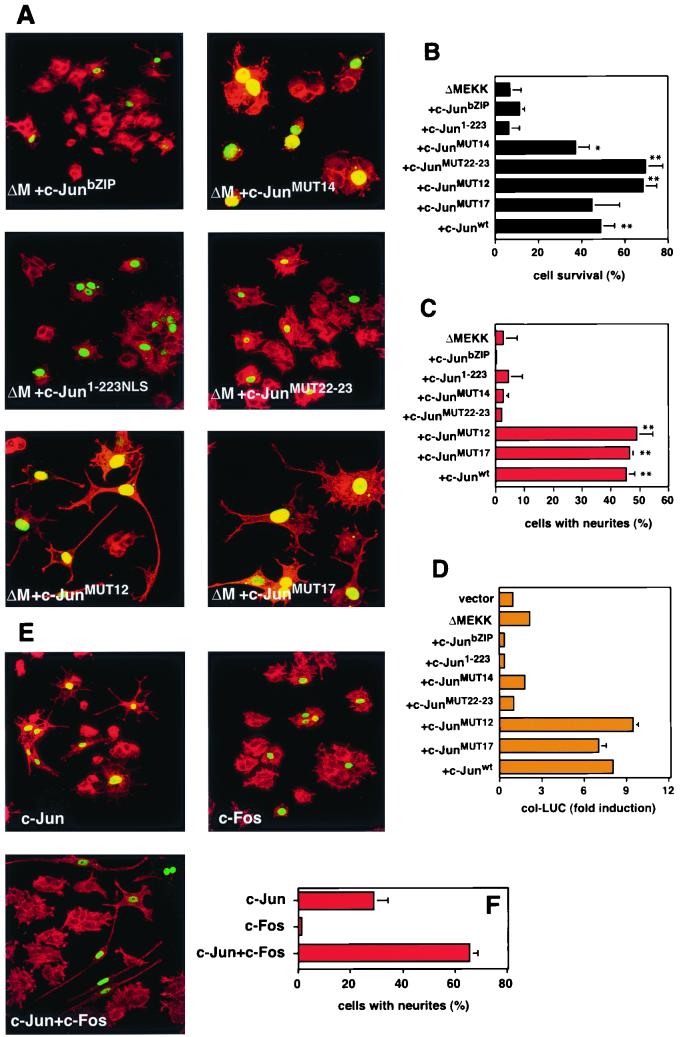
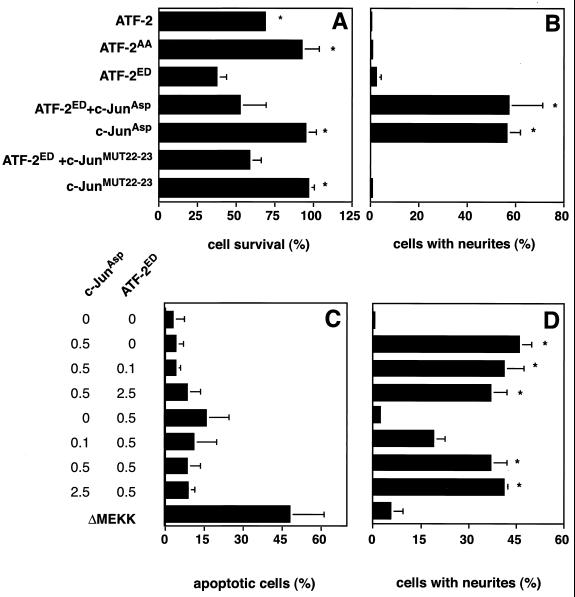
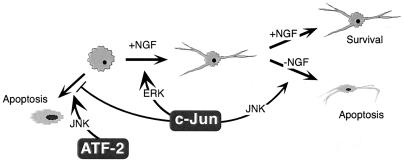
c-Jun activation by mitogen-activated protein kinases has been implicated in various cellular signal responses. We investigated how JNK and c-Jun contribute to neuronal differentiation, cell survival, and apoptosis. In differentiated PC12 cells, JNK signaling can induce apoptosis and c-Jun mediates this response. In contrast, we show that in PC12 cells that are not yet differentiated, the AP-1 family member ATF-2 and not c-Jun acts as an executor of apoptosis. In this context c-Jun expression protects against apoptosis and triggers neurite formation. Thus, c-Jun has opposite functions before and after neuronal differentiation. These findings suggest a model in which the balance between ATF-2 and Jun activity in PC12 cells governs the choice between differentiation towards a neuronal fate and an apoptotic program. Further analysis of c-Jun mutants showed that the differentiation response requires functional dimerization and DNA-binding domains and that it is stimulated by phosphorylation in the transactivation domain. In contrast, c-Jun mutants incompetent for DNA binding or dimerization and also mutants lacking JNK binding and phosphorylation sites that cannot elicit neuronal differentiation efficiently protect PC12 cells from apoptosis. Hence, the protective role of c-Jun appears to be mediated by an unconventional mechanism that is separable from its function as a classical AP-1 transcription factor.
Figures
References
-
- Behrens A, Sibilia M, Wagner E F. Amino-terminal phosphorylation of c-Jun regulates stress-induced apoptosis and cellular proliferation. Nat Genet. 1999;21:326–329. - PubMed
-
- Bohmann D, Tjian R. Biochemical analysis of transcriptional activation by Jun: differential activity of c- and v-Jun. Cell. 1989;59:709–717. - PubMed
-
- Bos T J, Monteclaro F S, Mitsunobu F, Ball A R, Chang C H W, Nishimura T, Vogt P K. Efficient transformation of chicken embryo fibroblasts by c-Jun requires structural modifications in coding and noncoding sequences. Genes Dev. 1990;4:1677–1687. - PubMed
-
- Davis R J. Signal transduction by the JNK group of MAP kinases. Cell. 2000;103:239–252. - PubMed
-
- Dérijard B, Hibi M, Wu I, Barrett T, Su B, Deng T, Karin M, Davis R. JNK1: a protein kinase stimulated by UV-light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell. 1994;76:1025–1037. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous