

Corneal collagen fibril structure in three dimensions: Structural insights into fibril assembly, mechanical properties, and tissue organization
- PMID: 11390960
- PMCID: PMC34664
- DOI: 10.1073/pnas.111150598
Corneal collagen fibril structure in three dimensions: Structural insights into fibril assembly, mechanical properties, and tissue organization
Abstract
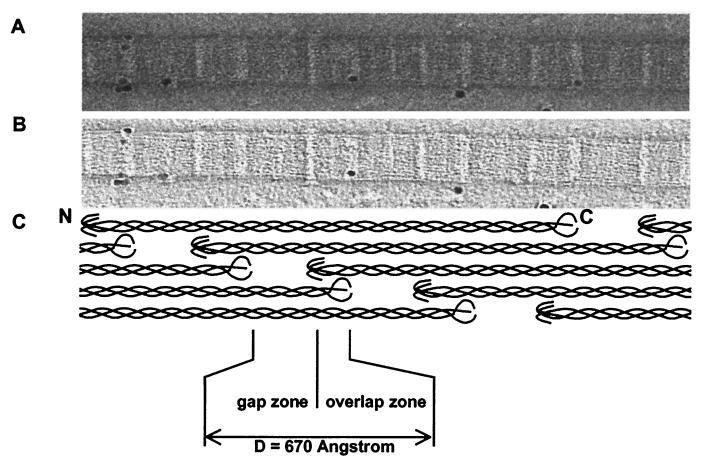
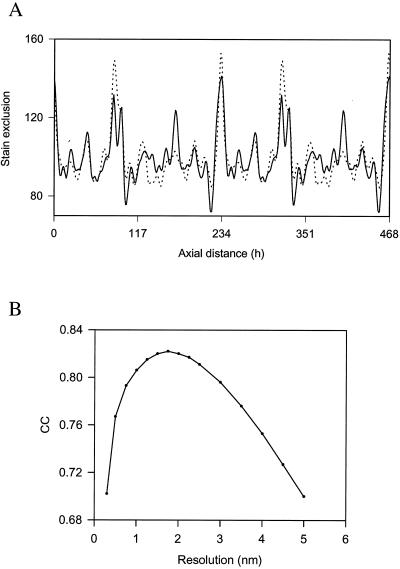
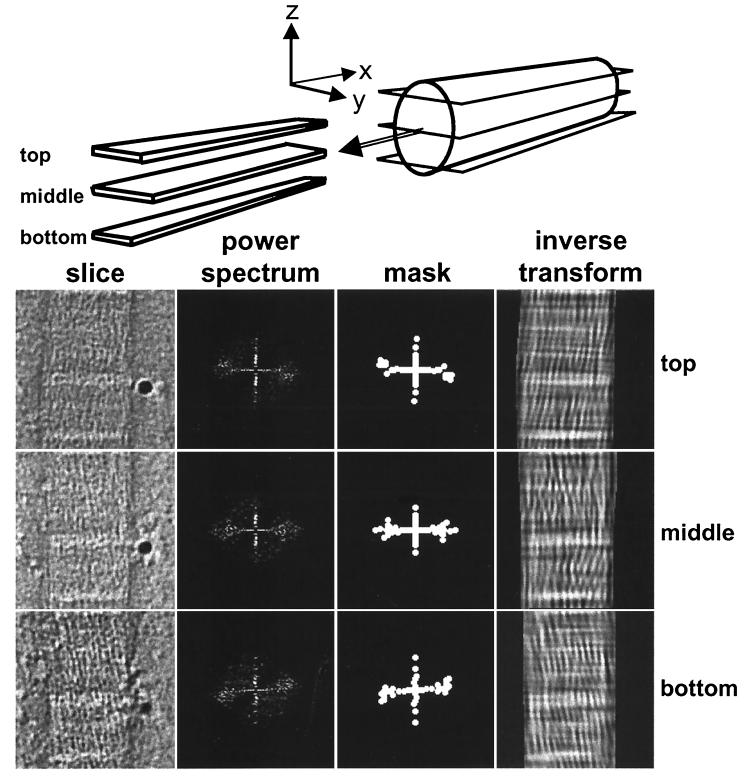
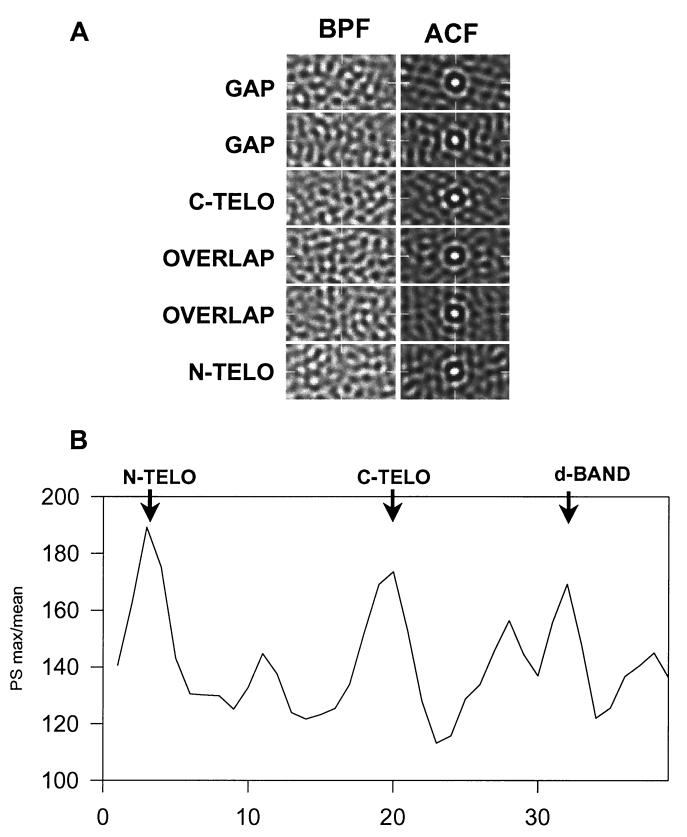
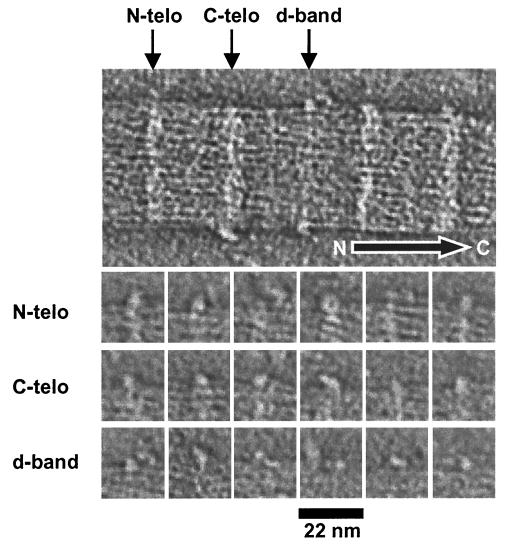
The ability of the cornea to transmit light while being mechanically resilient is directly attributable to the formation of an extracellular matrix containing orthogonal sheets of collagen fibrils. The detailed structure of the fibrils and how this structure underpins the mechanical properties and organization of the cornea is understood poorly. In this study, we used automated electron tomography to study the three-dimensional organization of molecules in corneal collagen fibrils. The reconstructions show that the collagen molecules in the 36-nm diameter collagen fibrils are organized into microfibrils (approximately 4-nm diameter) that are tilted by approximately 15 degrees to the fibril long axis in a right-handed helix. An unexpected finding was that the microfibrils exhibit a constant-tilt angle independent of radial position within the fibril. This feature suggests that microfibrils in concentric layers are not always parallel to each other and cannot retain the same neighbors between layers. Analysis of the lateral structure shows that the microfibrils exhibit regions of order and disorder within the 67-nm axial repeat of collagen fibrils. Furthermore, the microfibrils are ordered at three specific regions of the axial repeat of collagen fibrils that correspond to the N- and C-telopeptides and the d-band of the gap zone. The reconstructions also show macromolecules binding to the fibril surface at sites that correspond precisely to where the microfibrils are most orderly.
Figures
References
-
- Wess T J, Hammersley A P, Wess L, Miller A. J Struct Biol. 1998;122:92–100. - PubMed
-
- Eikenberry E F, Childs B, Sheren S B, Parry D A, Craig A S, Brodsky B. J Mol Biol. 1984;176:261–277. - PubMed
-
- Hulmes D J S, Miller A. Nature (London) 1979;282:878–880. - PubMed
-
- Fraser R D B, MacRae T P, Miller A. J Mol Biol. 1987;193:115–125. - PubMed
-
- Miller A, Tocchetti D. Int J Biol Macromol. 1981;3:9–18.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
