

Kinetic proofreading models for cell signaling predict ways to escape kinetic proofreading
- PMID: 11390967
- PMCID: PMC34662
- DOI: 10.1073/pnas.121172298
Kinetic proofreading models for cell signaling predict ways to escape kinetic proofreading
Abstract
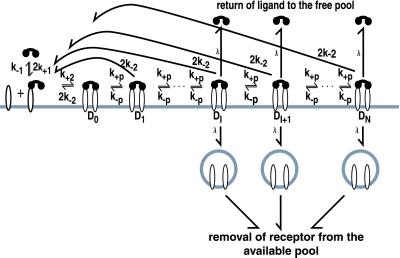
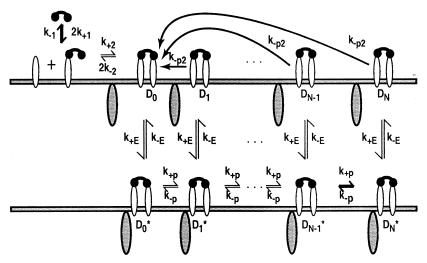
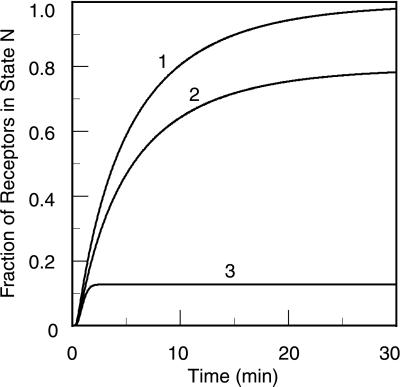
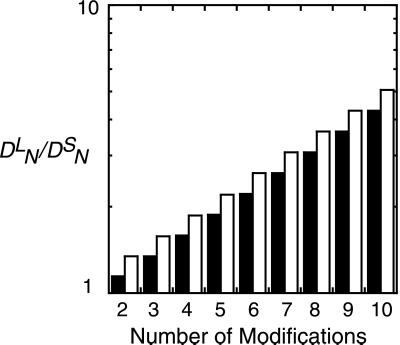
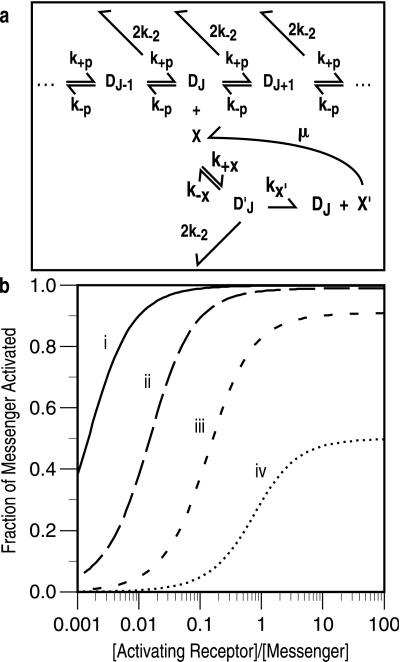
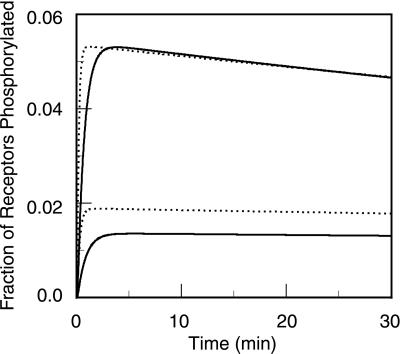
In the context of cell signaling, kinetic proofreading was introduced to explain how cells can discriminate among ligands based on a kinetic parameter, the ligand-receptor dissociation rate constant. In the kinetic proofreading model of cell signaling, responses occur only when a bound receptor undergoes a complete series of modifications. If the ligand dissociates prematurely, the receptor returns to its basal state and signaling is frustrated. We extend the model to deal with systems where aggregation of receptors is essential to signal transduction, and present a version of the model for systems where signaling depends on an extrinsic kinase. We also investigate the kinetics of signaling molecules, "messengers," that are generated by aggregated receptors but do not remain associated with the receptor complex. We show that the extended model predicts modes of signaling that exhibit kinetic discrimination for some range of parameters but for other parameter values show little or no discrimination and thus escape kinetic proofreading. We compare model predictions with experimental data.
Figures
Comment in
-
Signaling cascades: escape from kinetic proofreading.Proc Natl Acad Sci U S A. 2001 Jun 19;98(13):6989-90. doi: 10.1073/pnas.131215498. Proc Natl Acad Sci U S A. 2001. PMID: 11416175 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
