

Jasmonate-dependent induction of indole glucosinolates in Arabidopsis by culture filtrates of the nonspecific pathogen Erwinia carotovora
- PMID: 11402212
- PMCID: PMC111174
- DOI: 10.1104/pp.126.2.849
Jasmonate-dependent induction of indole glucosinolates in Arabidopsis by culture filtrates of the nonspecific pathogen Erwinia carotovora
Abstract
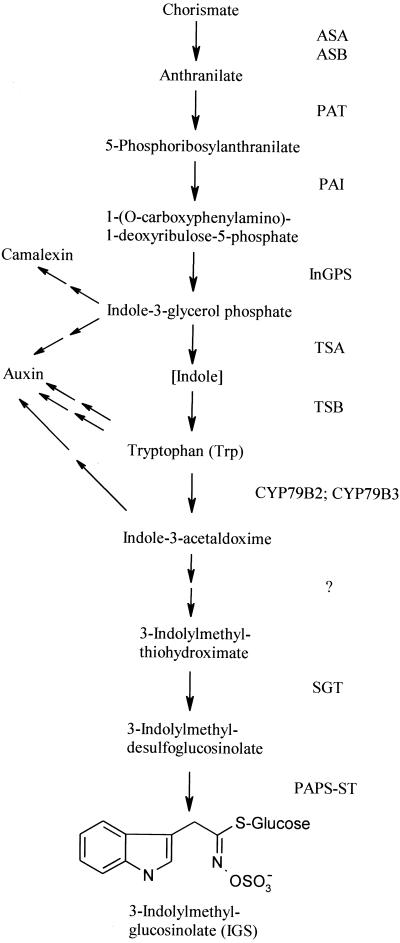
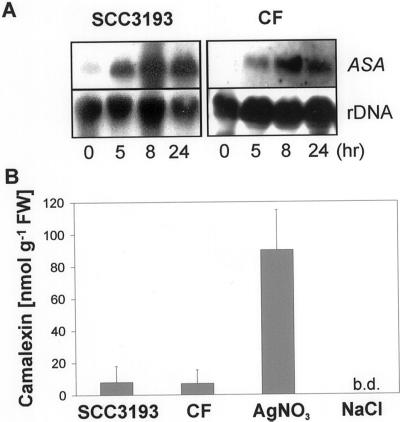
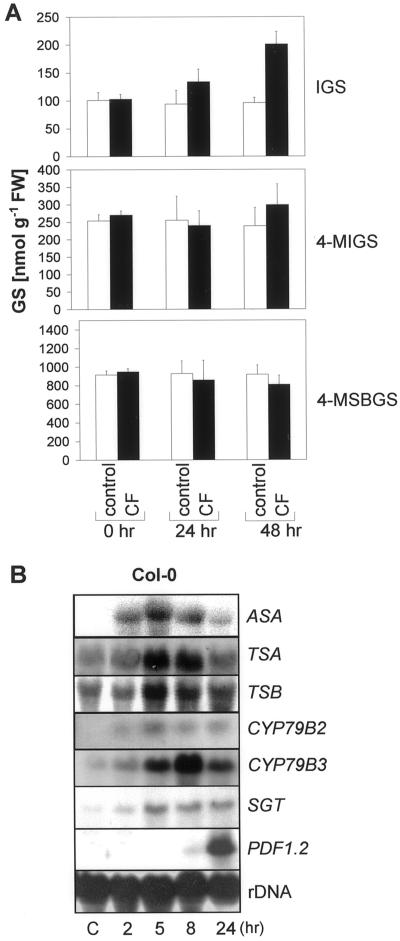
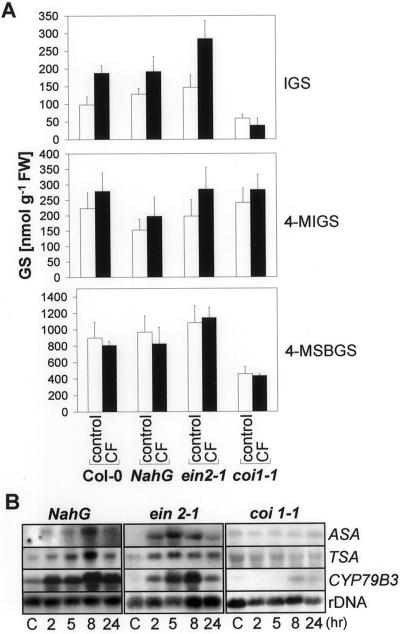
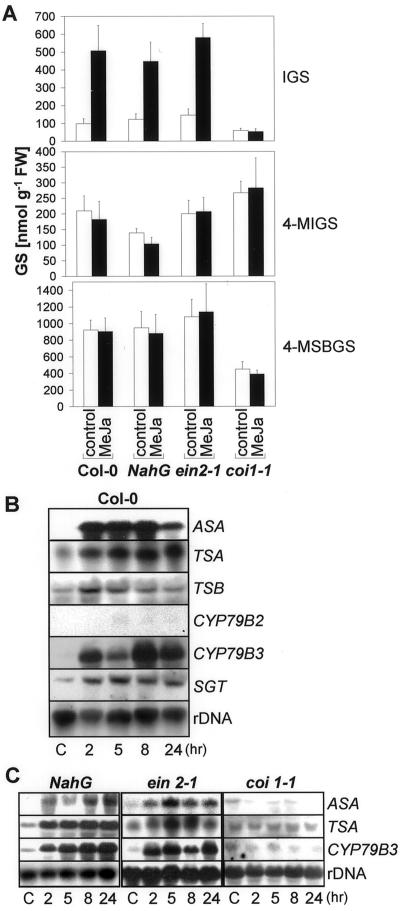
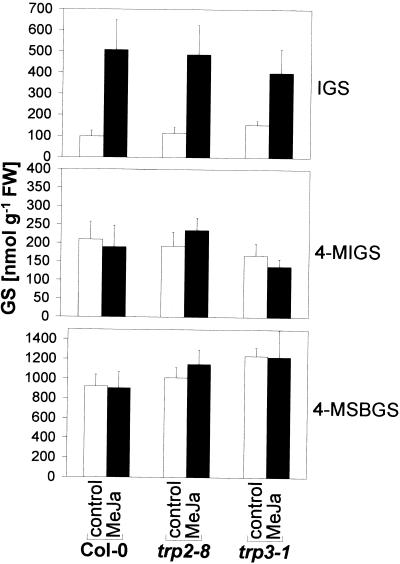
Elicitors from the plant pathogen Erwinia carotovora trigger coordinate induction of the tryptophan (Trp) biosynthesis pathway and Trp oxidizing genes in Arabidopsis. To elucidate the biological role of such pathogen-induced activation we characterized the production of secondary defense metabolites such as camalexin and indole glucosinolates derived from precursors of this pathway. Elicitor induction was followed by a specific increase in 3-indolylmethylglucosinolate (IGS) content, but only a barely detectable accumulation of the indole-derived phytoalexin camalexin. The response is mediated by jasmonic acid as shown by lack of IGS induction in the jasmonate-insensitive mutant coi1-1. In accordance with this, methyl jasmonate was able to trigger IGS accumulation in Arabidopsis. In contrast, ethylene and salicylic acid seem to play a minor role in the response. They did not trigger alterations in IGS levels, and methyl jasmonate- or elicitor-induced IGS accumulation in NahG and ethylene-insensitive ein2-1 mutant plants was similar as in the wild type. The breakdown products of IGS and other glucosinolates were able to inhibit growth of E. carotovora. The results suggest that IGS is of importance in the defense against bacterial pathogens.
Figures
References
-
- Agerbirk N, Bjergegaard C, Olsen CE, Sørensen H. Kinetic investigation of the transformations of indol-3-ylcarbinol into oligomeric indolyl compounds based on micellar elektrokinetic capillary chromatography. J Chromatography A. 1996;745:239–248.
-
- Agerbirk N, Olsen CE, Sørensen H. Initial and final products, nitriles, and ascorbigens produced in myrosinase-catalyzed hydrolysis of indole glucosinolates. J Agric Food Chem. 1998;46:1563–1571.
-
- Berger S, Bell E, Sadka A, Mullet JE. Arabidopsis thaliana Atvsp is homologous to soybean VspA and VspB, genes encoding vegetative storage protein acid phosphatases, and is regulated similarly by methyl jasmonate, wounding, sugars, light and phosphate. Plant Mol Biol. 1995;27:933–942. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
