

UV-damaged DNA-binding protein in the TFTC complex links DNA damage recognition to nucleosome acetylation
- PMID: 11406595
- PMCID: PMC150203
- DOI: 10.1093/emboj/20.12.3187
UV-damaged DNA-binding protein in the TFTC complex links DNA damage recognition to nucleosome acetylation
Abstract
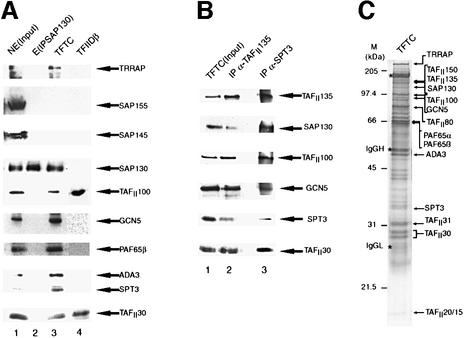
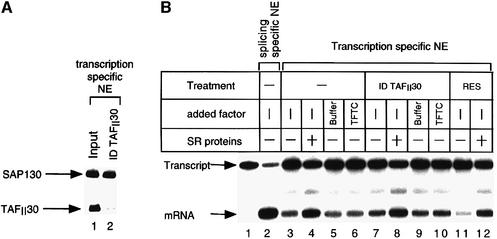
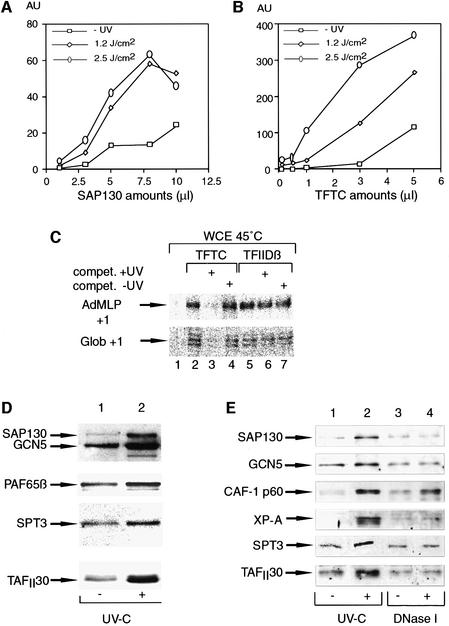
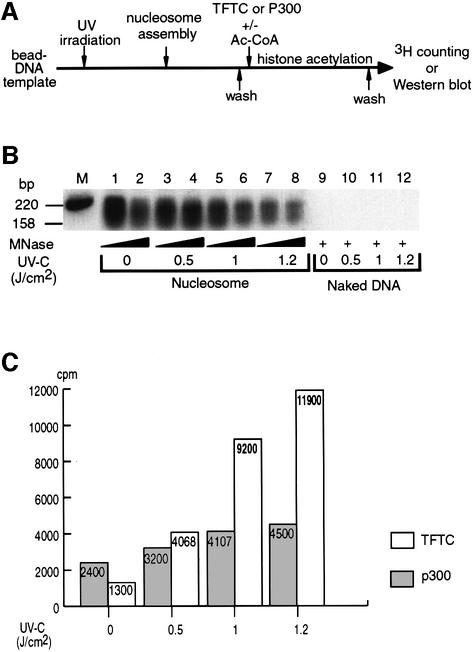
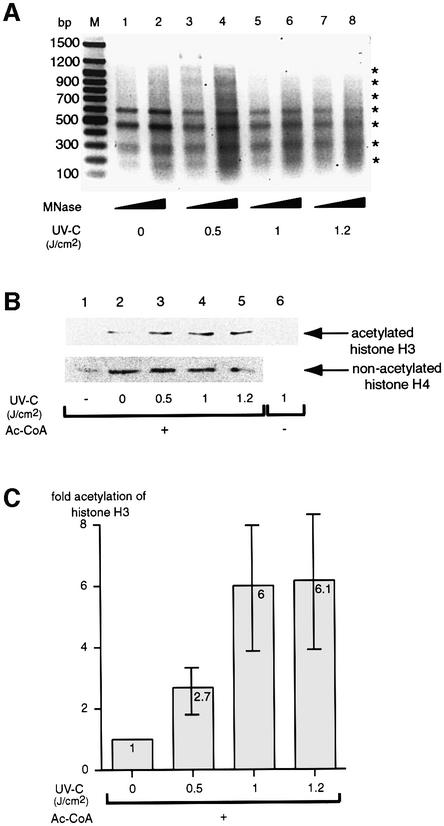
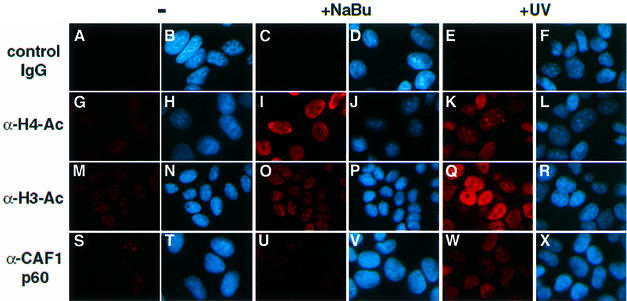
Initiation of transcription of protein-encoding genes by RNA polymerase II (Pol II) was thought to require transcription factor TFIID, a complex comprised of the TATA box-binding protein (TBP) and TBP-associated factors (TAF(II)s). In the presence of TBP-free TAF(II) complex (TFTC), initiation of Pol II transcription can occur in the absence of TFIID. TFTC containing the GCN5 acetyltransferase acetylates histone H3 in a nucleosomal context. We have identified a 130 kDa subunit of TFTC (SAP130) that shares homology with the large subunit of UV-damaged DNA-binding factor. TFTC preferentially binds UV-irradiated DNA, UV-damaged DNA inhibits TFTC-mediated Pol II transcription and TFTC is recruited in parallel with the nucleotide excision repair protein XP-A to UV-damaged DNA. TFTC preferentially acetylates histone H3 in nucleosomes assembled on UV-damaged DNA. In agreement with this, strong histone H3 acetylation occurs in intact cells after UV irradiation. These results suggest that the access of DNA repair machinery to lesions within chromatin may be facilitated by TFTC via covalent modification of chromatin. Thus, our experiments reveal a molecular link between DNA damage recognition and chromatin modification.
Figures
References
-
- Andel F., Ladurner,A.G., Inouye,C., Tjian,R. and Nogales,E. (1999) Three-dimensional structure of the human TFIID-IIA-IIB complex. Science, 286, 2153–2156. - PubMed
-
- Bell B. and Tora,L. (1999) Regulation of gene expression by multiple forms of TFIID and other novel TAFII-containing complexes. Exp. Cell Res., 246, 11–19. - PubMed
-
- Boggs B.A., Connors,B., Sobel,R.E., Chinault,A.C. and Allis,C.D. (1996) Reduced levels of histone H3 acetylation on the inactive X chromosome in human females. Chromosoma, 105, 303–309. - PubMed
-
- Bootsma D. and Hoeijmakers,J.H. (1993) DNA repair. Engagement with transcription. Nature, 363, 114–115. - PubMed
-
- Brand M., Leurent,C., Mallouh,V., Tora,L. and Schultz,P. (1999a) Three-dimensional structures of the TAFII-containing complexes TFIID and TFTC. Science, 286, 2151–2153. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
